Southern forms of Chiffchaff Phylloscopus collybita : observations from Iran and Armenia. |

Plate 1.
© A. R. Dean
Common Chiffchaff Phylloscopus collybita (probably menzbieri - see text), Kelardasht, N. Iran, April 2017.
Despite the advances afforded by molecular genetics, taxonomy remains an inexact science. Opinions vary, not only on species and subspecies boundaries but over the appropriate criteria to be employed in reaching taxonomic decisions. This situation is nowhere better exemplified than in the Chiffchaff complex. The BOU now follows the IOC World Bird List which recognises six subspecies of Common Chiffchaff Phylloscopus collybita viz. P. c. collybita, P. c. abietinus and P. c. tristis in the northern part of the species’ range and P. c. brevirostris, P. c. caucasicus and P. c. menzbieri farther south (Figures 1 & 2). Hereafter, each taxon - including also (Caucasian) Mountain Chiffchaff P. sindianus lorenzii - is generally referred to succinctly by its subspecific name.
Based upon mitochondrial DNA (mtDNA), morphology and vocalisations, Helbig, Seibold, Henning, Schottler & Wink (1996) and Clement, Helbig & Small (1998) greatly clarified understanding of the taxonomy of the Chiffchaff complex. However, the distinguishing features of the southern forms of Common Chiffchaff, brevirostris, caucasicus and menzbieri, remain far from firmly established, while Helbig et al. found the forms brevirostris and caucasicus to be virtually indistinguishable genetically. All three southern taxa have been described as close in appearance to abietinus (e.g. Clement et al. 1998). Other accounts suggest that, as a group, the southern forms are somewhat intermediate between abietinus and lorenzii but closer to the former, being rather 'browner' than typical abietinus and with reduced yet still-evident olive and yellow hues. However, references to menzbieri frequently acknowledge the lack of current information and an absence of photographs. The songs of the southern subspecies are all variants on the 'chiff-chaff' theme but pace and timbre vary while sonograms reveal distinct note structures. Counter-intuitively, the calls of the southern forms do not match the rising 'huit' call of collybita and abietinus but are very similar to the 'straight', plaintive 'eeep' call of tristis. Of the Chiffchaff complex as a whole, Helbig et al. wrote : '... vocal differences, like mitochondrial DNA sequences, reflect evolutionary differentiation within the chiffchaff complex over long time spans much better than does morphology, which is relatively uniform'.
In general, comments on the appearance of menzbieri presented in European literature are inconsistent and third-hand. As the form caucasicus was not described by Loskot until 1991, earlier historical accounts of menzbieri do not mention, yet alone compare, caucasicus. Presumably on the basis of a deficit of olive and yellow, Chiffchaffs from NE Iran to SW Transcaspia (i.e. the range associated with menzbieri) were included in tristis (or ‘fulvescens’) by Vaurie (1959), Williamson (1962) and Cramp et al. (1992), though the last added that it might belong with abietinus. Shirihai & Svensson (2018) also suggested plumage convergence between menzbieri and both 'fulvescens' and eastern abietinus. On distributional grounds alone, that Chiffchaffs from Kopet Dagh and NE Iran belong with tristis seems implausible.
Unfortunately, all but two of the original type specimens of menzberi were destroyed in an earthquake and fire at Ashkhabad but Loskot (2002) provided useful comments from the type description of Shestoperov (1937), including that the upperparts were 'olive grey-brownish with barely visible greenish touch' and the underparts 'whitish, especially pure white in the middle of belly and on under tail coverts', differing from abietinus 'mainly in an absence of yellow streaks on the underparts' and from fulvescens 'in the more greyish upperparts'. Two surviving specimens from the type series had been brought to ZMMU by Dementiev, who had examined the type series and noted that their upperparts were more greyish than in abietinus (Dementiev 1948). Loskot designated one as the lectotype, commenting that it was ‘.. in good condition and has all characters mentioned in the original description of Ph. c. menzbieri’. However, the paralectotype he concluded was not attributable to menzbieri and was probably a migrant of the ‘fulvescens’ type.
One of the very few accounts based upon first-hand observations of menzbieri in the Kopet Dagh, from where the original 'type specimens' were obtained, is that of Marova & Leonovich (1997). They noted that this region is rarely visited by ornithologists. Discussing distribution, the authors noted that Kopet Dagh was the established range but they included citations to menzbieri in the Elborz mountains of northern Iran. They suggested that the Elborz population was probably menzbieri but acknowledged that further information was required. This account provided explicit comments on the comparative appearance of menzbieri and caucasicus. It is in Russian but includes an English summary, from which the following is an extract (verbatim):
'Contrary to Ph. c. abietinus, Ph. c. menzbieri has only a trace of yellow colour on the breast, white belly and white undertail coverts. From Ph. c. caucasicus it differs both in the bright yellow colour of the axillars and the wrist and in the brighter green colour on the back and on the edge of the primaries and tail feathers'. Once again, rather white underparts enter the frame for the southern races but that menzbieri exhibits more 'green' on the upperparts than caucasicus seems unexpected, given the comments of Loskot and the details of the type description of Shestoperov that the 'greenish touch' on the upperparts of menzbieri was 'barely visible'. However, Irina Marova (in litt.) states that, while the appearance of menzbieri has little in common with tristis or lorenzii, the differences between menzbieri and caucasicus noted by Marova & Leonovich (1997) are relatively small.
Watson (1962), in his paper recommending reinstatement of the name brevirostris for the Chiffchaffs breeding in northern Turkey, provided a table of comparative characters of brevirostris, abietinus, menzbieri and lorenzii.. The distinguishing features included in the table are very rudimentary but the word 'brown' is allocated only to the upperparts of lorenzii. The description for the upperparts of abietinus is 'greyish green' and for brevirostris 'greenish grey' (implying that brevirostris is the greyer form). Watson described the upperparts of menzbieri as 'mostly grey, tinged greenish' [my emphasis] but it seems unlikely that this was based upon examination of extensive original material. He noted a west-to-east reduction in lipochrome pigment (as in the northern forms), resulting in whiter underparts, with reduced yellow streaking. The underparts of menzbieri he described simply as: 'white, no yellow'.
More recently, Copete & Lopéz (2013) and Shirihai & Svensson (2018) have questioned aspects of the prevailing taxonomy of Common Chiffchaff. Both sets of authors suggested, on the basis of morphology, that brevirostris be subsumed with collybita. Copete & Lopéz noted that photos of type specimens of caucasicus provided to them by Vladimir Loskot exhibited upperparts similar to abietinus but 'yellow much more clearly present on the edge of the wing, lower parts clearly whiter'. The legs were also noted to appear densely black, more so even than in tristis. They considered that the material they had examined showed a consistent appearance but concluded that examination of a longer series was required to confirm the validity of caucasicus. Despite the near-identical mtDNA of brevirostris and caucasicus, Shirihai & Svensson (2018) - who advocated merging brevirostris with collybita - suggested that caucasicus be merged with abietinus. While retaining menzbieri, at least pro tem, Shirihai & Svensson also raised doubts over that taxon's validity. This comes close to negating the entire group of southern subspecies. As the appearance of typical abietinus is also emphasised as very close to that of nominate collybita (e.g. Svensson 1992), functional differences in appearance among the Common Chiffchaff races are inferred to be very subtle indeed (with a more-singular image emerging only for 'classic' tristis). In contrast with other subspecies, Copete & Lopéz (2013) concluded that they could not comment on the morphology of menzbieri, as there was a lack of accessible data.
The following text is based upon photographs and sound-recordings of Common Chiffchaffs observed at Kelardasht and Kiaser in the Elborz Mountains of N. Iran during April 2017 and in the Armenian Caucasus at Dilijan and the Vorotan Valley during May 2018. At that time, a clear image of the precise distributions, the appearances of, and distinctions between, the southern races were not forthcoming from the existing literature. Figure 1 shows the breeding ranges of the southern species and subspecies as depicted by Copete & López (2013) and modified from Handbuch der Voegel Mitteleuropas, Glutz von Blotzheim & Bauer (1991). Of necessity, the ranges shown were no more than approximate, as the full distributions of the southern subspecies of Common Chiffchaff remained imprecisely clarified (while (Caucasian) Mountain Chiffchaff (lorenzii) certainly has a wider distribution than indicated in Figure 1, e.g. extending into the western Elborz Mountains of Iran). Superimposed on this map are the approximate locations of Kelardasht and Kiaser, in Iran, and Dilijan and Goris in Armenia.
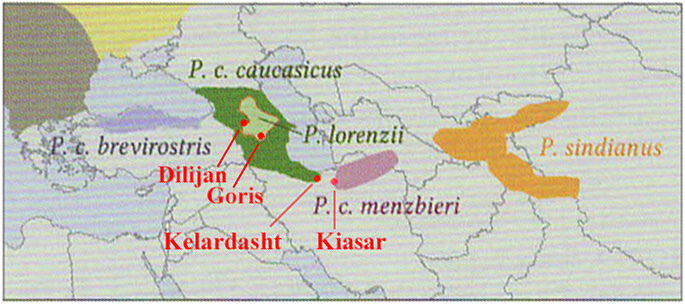
Figure 1.
The locations of Kelardasht and Kiasar, in Iran and
Dilijan and Goris in Armenia, superimposed on
range map for caucasicus and menzbieri (and other southern forms)
as portrayed by Copete & López (2013)
and modified from Handbuch der Voegel Mitteleuropas, Glutz von Blotzheim
& Bauer (1991).
Kelardasht is at the eastern extreme of the range depicted for caucasicus in Copete & López (2013) while Kiasar lies close to the western end of the range depicted for menzbieri (its core range regarded as Kopet Dagh in Turkmenistan) - see Fig. 1 above. As at April 2017, how far west menzbieri extends was not established. Similarly vague was the true extent of the range of caucasicus along the southern shores of the Caspian Sea. On the basis of range, it was not possible in April 2017 to allocate the Chiffchaffs at Kelardasht and Kiasar to one form or the other. The Armenian birds were equated with caucasicus but those in Iran were left indeterminate between caucasicus or menzbieri, as the distinguishing features and precise ranges of these two forms were inadequately defined.
Subsequently, a more-comprehensive genetic study of the Chiffchaff complex was published by Raković et al. (2019). This is essentially a genetic study and did not examine morphology or vocalisations. Its findings broadly support the conclusions of Helbig et al. (1996) and the validity of the currently recognized subspecies of Common Chiffchaff, while also describing the genetic makeup among the newly discovered Chiffchaff population breeding at Mt. Hermon, on the border between Lebanon and Syria - see Raković et al. for details. Although Chiffchaffs from the range of brevirostris within Turkey were not sampled, individuals with the ‘caucasicus’ ND2 sequence were found at sampled locations (such as the Balkans and Mt. Hermon) that are much closer to the known range of brevirostris and this was consistent with the assignation of this clade to a combined ‘P. c. brevirostris/caucasicus’ (Helbig et al. (1996) having found these two to be indistinguishable on the basis of mtDNA). Significantly, a separate clade was identified around Transcaucasia, south and east from, and genetically distinct from, the brevirostris / caucasicus clade. The inference that this clade equated with menzbieri (the only recognised subspecies south and east of 'caucasicus') was supported by the single genetic sample of menzbieri in GenBank. Thus, on geographic and genetic grounds, this Transcaucasus clade was reasonably equated with menzbieri. The authors note that in some taxonomic publications the form(s) of Common Chiffchaff found in southernmost Armenia and northern Iran have been left undefined (e.g. in the Checklist of Birds of the World compiled by Clements et al. 2015) and, to place this new interpretation beyond doubt, a much larger and representative series of genetic material was required from across N Iran and from Kopet Dagh. The implication, however, is that menzbieri extends from southern Armenia (south of Goris), continuously through the Hyrcanian forest of the Elborz mountains of N. Iran and east to Kopet Dagh in Turkmenistan, a much more extensive range than the often-quoted 'isolated range' around Kopet Dagh.
Figure 2 below shows the ranges of the Chiffchaff complex as depicted by Raković et al. in 2019, based upon genetic clades.
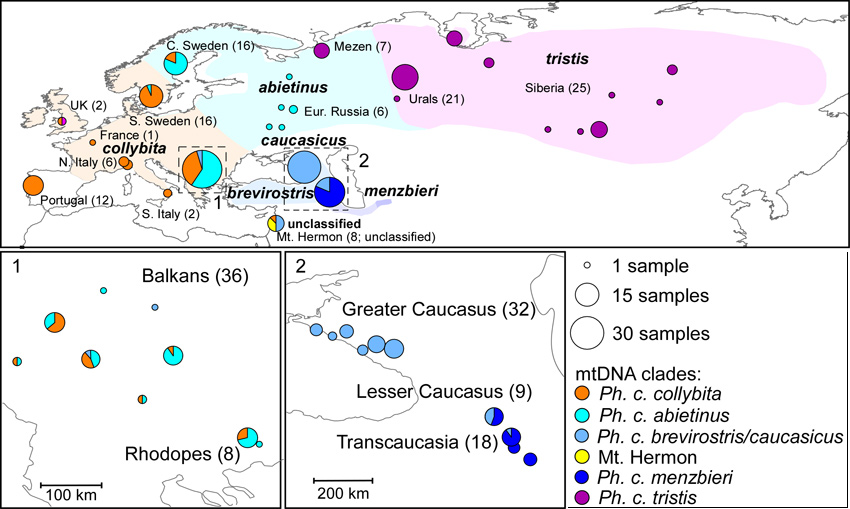
Figure 2.
'Ranges of Common Chiffchaff subspecies, sample localities
and mtDNA clades', from
Raković et al. (2019).
Given the validity of Raković et al.'s conclusions, the Iranian Elborz birds encountered in April 2017 would be menzbieri; those in May 2018 at Dilijan in Armenia were in the region where caucasicus and menzbieri overlap, while those in the Vorotan Valley near Goris were at the northern limit of the range of menzbieri where slight introgression of genes is still possible (see Figure 2). Hopefully, the photos and sonograms below, from the Iranian Elborz Mountains and the Armenian Lesser Caucasus, will provide useful supplementary (and relatively 'accessible') reference material on the appearance and vocalisations of the enigmatic Chiffchaff forms inhabiting these areas.
Plates 2 to 6 show Common Chiffchaffs Phylloscopus collybita observed in the central Elborz Mountains near Kelardasht, in northern Iran, during April 2017. The species was quite numerous in light woodland (including oak, hornbeam and hawthorn) at an altitude of around 1500m, a habitat shared with Green Warblers P. nitidus. Chiffchaffs of similar appearance and vocalisations were encountered subsequently in denser woodlands farther east, near Kiasar.

Plate 2
© Ali Alieslam

Plate 3
© Ali Alieslam

Plate 4
© Steve Rooke

Plate 5
© Rob Tizard

Plate 6
© Rob Tizard
Plates 2 to 6. Common Chiffchaffs, c. 1500m in Elborz Mountains near Kelardasht, Iran, April 2017
The appearance of the Elborz Chiffchaffs differed in several respects from that of typical abietinus. Their upperparts were greyish-brown (greyer on the mantle, somewhat browner on the crown). Their underparts were conspicuously white (though slightly sullied), lacking field-evident yellow anywhere in the body plumage (though with a tinge at the front of the upper eye-ring). The upperparts away from the crown were diffusely and inconspicuously streaked with olive and there were well-developed olive-yellow fringes to the remiges and rectrices. A pale buff supercilium was shallow but rather long, extending well behind the eye. The legs and feet were decidedly black while there was a conspicuous pale base to the lower mandible. Often the plumage appeared rather pale overall but, at close range and in duller light, the browner aspect to the upperparts was more apparent. A deficit of olive and yellow in the body plumage but combined with olive-yellow fringes to the flight feathers is no doubt the source of analogies which have been made with 'fulvescens', with which menzbieri was included historically by Vaurie (1954) and Williamson (1962), despite their distributions being separated by over 1500km (see Appendix). There is a suggestion of 'fulvescens' in the appearance of the upperparts but the Iranian chiffchaffs have more conspicuous yellowish-olive fringes to the wing-feathers and tail-feathers, greyer-tinged upperparts and much whiter underparts (lacking the strong buff wash on the flanks of tristis s.l.). Viewed in profile (e.g. Plate 2) or from below (e.g. Plate 1), the 'whiteness' of their underparts is consistently distinctive.
The precise colour-balance in digital images is influenced by light-conditions, camera settings and even the make and model of the camera (see https://deanar.org.uk/tristis/photoissues.htm). Inevitably, this applies too in the photos above but, as a set, they illustrate well the colour nuances which distinguish the Elborz Chiffchaffs from typical abietinus and I am grateful to Ali Alieslam, Steve Rooke and Rob Tizard for providing their excellent images.
At higher altitudes above 2500m, on relatively bare mountain slopes with scrub and isolated clumps of small trees, these grey-tinged chiffchaffs with whitish underparts were replaced by (Caucasian) Mountain Chiffchaffs P. lorenzii which, even in the brightest light, exhibited much more densely brown upperparts (lacking olive), a distinctive fulvous-buff wash on the breast, 'plain' wing and tail edges (lacking the prominent yellow-olive fringes of the Common Chiffchaffs) and a more-prominent supercilium (together with subtly different vocalisations). Although exhibiting a more saturated brown hue to their upperparts, these lorenzii converged more with typical tristis than did the Common Chiffchaffs.
Thus, the appearance of the Elborz Chiffchaffs conformed well with key features attributed to menzbieri by Shestoperov (1937), Marova & Leonovich (1997) and Loskot (2002). Above all, the predominantly white appearance of their underparts is distinctive. It is likely that a minority of abietinus from the east of its range exhibits a relatively grey and white appearance, which may converge with that of the Iranian Chiffchaffs. Given that the Iranian Chiffchaffs are now attributable to menzbieri, then this convergence no doubt underpins suggestions that menzbieri be subsumed with abietinus (e.g. Svensson (1992), though Shirihai & Svensson (2018) provisionally retained menzbieri pending further research). However, the majority of abietinus are very similar to collybita (as emphasised by Svensson 1992 and Shirihai & Svensson 2018). Conversely, the greyer aspects in the upperparts of menzbieri and, especially, the whiteness of much of their underparts, is a striking and consistent aspect of the Elborz and the Caucasian birds. Together with its tristis-like calls and genetic divergence, menzbieri seems deserving of taxonomic recognition.
For discussion of plumage variability in abietinus and the identity of 'greyer' Chiffchaffs encountered occasionally in the UK and western Europe see: https://deanar.org.uk/tristis/tristis.htm#plumagelimits and https://deanar.org.uk/tristis/tristis.htm#greyChiffs.
In May 2018, Common Chiffchaffs were observed, photographed and sound recorded in the Dilijan region and in the Vorotan Valley of Armenia. Again, Green Warblers were found in the same habitats while lorenzii overlapped in altitudinal range. At the time these Armenian birds were assumed to be caucasicus but the recent genetic studies by Raković et al. indicate that menzbieri extends into Armenia south of Goris and that Transcaucasus and especially the Lesser Caucasus comprise a transition zone between caucasicus and menzbieri (see Figure 2). Thus, the Chiffchaffs at Dilijan were close to the border between caucasicus and menzbieri, with the genetic samples analysed by Raković et al. comparably distributed between the two (4 and 5 respectively). Those in the Vorotan Valley near Goris were close to the northern extremity of menzbieri but where there is still slight introgression from caucasicus (see Figure 2).

Plate 7
© A. R. Dean
.
Plate 8
© A. R. Dean

Plate 9
© A. R. Dean
Plates 7 to 9. Common Chiffchaffs, near Dilijan, Armenia, May 2018.
This area is in the region where caucasicus overlaps with menzbieri,
if following the
conclusions
of
Raković et al. (2019).
The Chiffchaffs in Armenia in
plates 7 to 9 share rather white underparts with the Iranian individuals but with
olive more apparent on the upperparts. This perhaps implies that
caucasicus is slightly more olive, not
less, than menzbieri (c.f. Marova & Leonovich 1997).

Plate 10
© A. R. Dean
Common Chiffchaff, near Dilijan, Armenia, May 2018. Close-up of wings showing contrasting yellowish-olive fringes to flight feathers.

Plate 11
© A. R. Dean
Common Chiffchaff, near Dilijan, Armenia, May 2018.
Close-up of underparts, with position providing soft shade
and avoiding
over-illumination or 'burn out'. The distinctive 'white' ground-colour to lower
breast and belly is thereby
confirmed.
The distinctly white-looking underparts, well-defined yellow-olive fringes to flight-feathers and 'densely black' legs accord with the impressions noted by Copete & López (2013), based upon photographs of type specimens of caucasicus (provided to them by Vladimir Loskot). However, they are also consistent with characters noted for menzbieri and how closely these features may be shared by the two forms remains to be resolved.

Plate 12
© A. R. Dean
Common Chiffchaff, Vorotan Valley near Goris, Armenia, May
2018. Chiffchaffs 'south of Goris' are part of the Transcaucasus clade,
equated
with menzbieri
by
Raković et al.
(2019) - see Figure 2. Thus, the individual in plate 12 is from a region
where menzbieri and caucasicus converge and there is some introgression.
As with the Chiffchaffs
in N. Iran
in plates 1 to 6, it exhibits little olive in the grey-tinged upperparts
and a deficit of yellow below, though there is some yellow near the bend of the
wing.
Helbig et al. (1996) and Clement et al. (1998) provided sonograms of the songs and calls of all subspecies. More recently, Marova et al. (2021) have published a comprehensive study of songs, including a multi-variate analysis. These are used to provide a platform by which to evaluate sound-recordings made in Iran in April 2017 and Armenia in May 2018.
A recording of the song from Kelardasht, Iran, in April 2017, can be heard <here>. Compared with collybita / abietinus, the notes have a harder, 'punchier' and more staccato delivery. A sonogram appears as the upper graph in Figure 3, with comparative sonograms of caucasicus and menzbieri from Helbig et al. presented below.
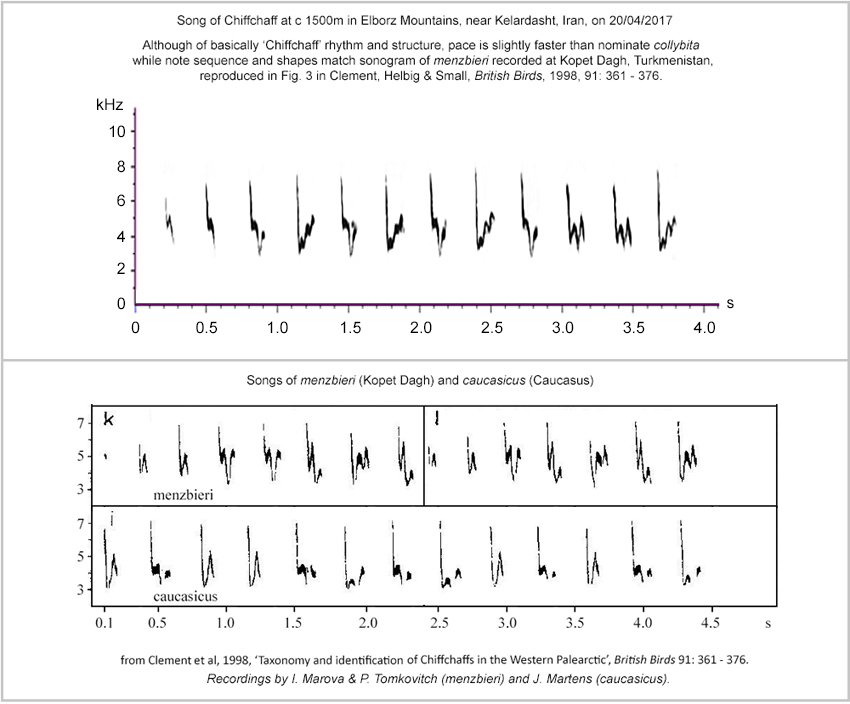
Figure 3.
Sonogram of Chiffchaff song recorded at Kelardasht, in Elborz Mts,
Iran, April 2017
compared with sonograms published by Helbig et al.(1996) and Clement
et al. (1998).
The sonogram from Kelardasht shows three introductory 'chiff' notes, simple and somewhat h-shaped notes. These also appear in the sonograms of caucasicus by J. Martens and menzbieri by I. Marova & P. Tomkovitch but those in the Kelardasht sonogram have a terminal component descending to a lower frequency. Subsequent notes are more complex but each note-structure in the Iranian recording from April 2017 has a close match in the sonograms of menzbieri presented by Helbig et al. (1996). Similar though less precise matches for many notes can be found in Helbig et al.'s sonogram of caucasicus.
Many of the notes in the sonograms of both of these taxa have analogs in sonograms of collybita and abietinus song. In the menzbieri sonograms, the exceptions are in the note found at the fourth, sixth, eighth and twelfth positions in the Iranian recording from 2017, position six in Helbig et al.'s first sonogram of menzbieri and positions five and seven in their second sonogram. This note is a 'chaff'-style note and includes a rising 'saw-tooth' flourish or appendix rising acutely from the base of the initial down-stroke. The peak frequency of this appendix reaches to between 40% and 60% of the peak (starting) frequency of the note overall. Such a sharply rising basal 'appendix' is not found in collybita or abietinus. This is the most 'emphatic' note, with a rather rounded vowel sound ('choiff' rather than 'chaff') and is very evident to the ear in the sound-recording from 2017. Together with overall note 'pacing' and ratio of 'chiffs' to 'chaffs', this 'rising sawtooth' note generates the different character from western Chiffchaff song.
A recording of the song from the Vorotan Valley, Armenia, can be heard <here> while a sonogram is provided in Figure 4.
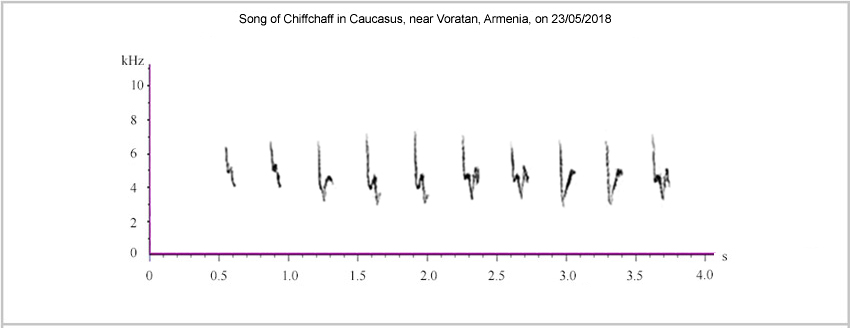
Figure 4.
Song of Chiffchaff in Vorotan Valley, Armenia, May 2018.
The recording from May 2018 in the Vorotan Valley of Armenia is from a region where introgression of caucasicus into menzbieri commences, according to the findings of Raković et al. (2019). Thus, some overlap might be anticipated in song-structure. The recording has a more even sound than the song recorded in Iran in April 2017, with most notes being 'chiff' style rather than 'chaff' style. However, sonograms share the simple introductory notes (with a pronounced descending terminal stroke). Also, subsequent notes include, at positions eight and nine, a note with a rising appendix positioned wholly above the base frequency of the initial downstroke, drawing comparisons to menzbieri. However, the rising appendix here appears relatively even, lacking a saw-tooth ('up-down, up-down') structure. Listening to the songs from Iran and Armenia 'back-to-back' <here>, the 'saw-tooth' note in the Iranian song is emphatic and contrastingly 'chaff' (or 'choiff') style while notes eight and nine (with the evenly rising appendix) in the Armenian recording are relatively soft and subdued.
This note, with an evenly rising appendix, is found in the sonogram for caucasicus provided by Helbig et al. whereas the note found in menzbieri (with a 'sawtooth' structure rising strongly from the minimum frequency point) is not present. Thus, in May 2018 and on the basis of these recordings, a 'sawtooth' structure to the appendix seemed to be associated with menzbieri while a straight structure was associated with caucasicus.
More-recently, however, a study has been published by Marova et al. (2021) that suggests a level of overlap between the forms in this respect. Marova et al. annotated a stylized version of a note with a saw-tooth appendix as characteristic of both menzbieri and caucasicus but rarely found in other taxa. A recording of a Chiffchaff at Zemo Teleti Hills, near Tibilisi in Georgia, in April 2022, was published on xeno-canto by Oscar Campbell (xc734545). This too shows some notes with a 'straight' rising appendix and others with a slightly saw tooth structure. The location is firmly within the breeding range of caucasicus but it was not established whether the singer was on territory. Inspections of the 'saw-tooth' notes in the caucasicus sonograms published by Marova et al. and the sonogram published by Campbell, do suggest that the 'saw-tooth' structures therein tend to be single and of more-limited frequency range ('less emphatic') than in the menzbieri recordings and it may be that a series of emphatic 'saw-tooth' notes with several well-developed 'teeth' may favour menzbieri but this requires confirmation from a much longer series of recordings of both taxa from the heartlands of their breeding ranges.
While many note-structures of the Iranian and Armenian birds are broadly similar, they differ significantly from the note-structures in the sonograms of brevirostris by P. S. Hansen from Ulu Dagh, Turkey, published by Helbig et al., and in a recording by Manuel Sanchez of brevirostris from Istanbul, available on xeno-canto as xc142271. The song of brevirostris is dominated by a single, repeated note with double or even triple 'spurs' or 'teeth' on a downward frequency scale, as in Figure 5.
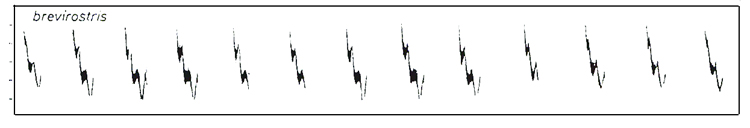
Figure 5. Sonogram of brevirostris song by P. S.
Hansen from Ulu Dagh, Turkey.
Reproduced from Helbig et al. (1996) & Clement et al.
(1998).
Thus, an appendix rising acutely and significantly from the base of the original downstroke appears indicative of one or other of menzbieri or caucasicus and is unlikely to be found in brevirostris, abietinus or collybita.
Marova et al. (2021) also categorised the songs of all Chiffchaff subspecies, and lorenzii too, using seven different measurements of frequency and modulation parameters. Disconcertingly, using this multi-variable approach, they found grouping between Armenian caucasicus and menzbieri but that menzbieri from Kopet Dagh differed from this group. They wrote: 'Two [bioacoustic] samples of caucasicus and two samples of menzbieri are clustered into one node; however, all three samples from the Caucasus, including the Armenian menzbieri, form a well-evidenced distinct node and appear to be separate from the Kopet Dagh’s menzbieri'. They did not discuss whether this could be a product of introgression in southern Armenia but it indicates the need for further investigation of the songs of menzbieri from Kopet Dagh and caucasicus from the Greater Caucasus, away from the convergence and overlap zones.
(Grishchenko et al. (2016) analysed the songs of a newly-established breeding population of Common Chiffchaffs in the Crimea. They concluded that certain elements in the song were shared with caucasicus but not with abietinus and concluded that the Crimean population originated from caucasicus. Using the sonograms published in 1996 by Helbig et al. (and Clement et al. in 1998) they also concluded that menzbieri (and brevirostris too) contained subspecifically distinct elements. Unfortunately, they did not elaborate or include illustrative, annotated sonograms of all forms).
A recording of the call of Chiffchaffs at Kelardasht, Iran, in April 2017 can be heard <here>.
A recording of the call of Chiffchaffs in the Vorotan Valley, Armenia, in May 2018 can be heard <here>.
Figure 6 provides sonograms of the calls recorded in Iran and Armenia while, for comparison, sonograms attributed to brevirostris, caucasicus, menzbieri and tristis are included, reproduced from Helbig et al (1996) and Clement et al. (1998).
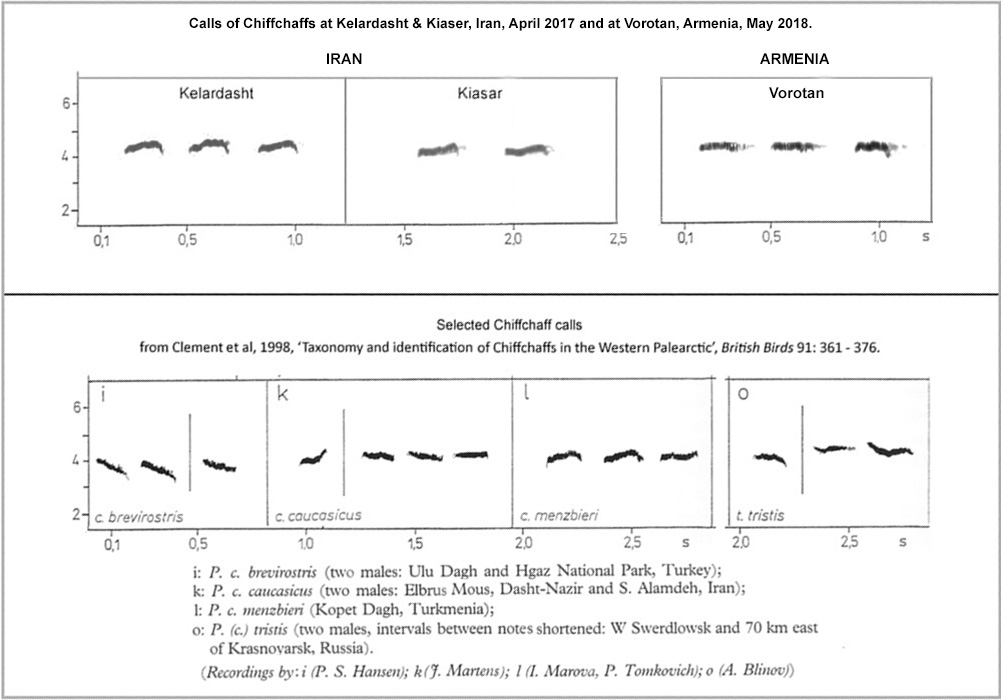
Figure 6.
Clearly, all these calls are relatively straight or 'flat', though brevirostris has a slightly descending frequency and the first call attributed to caucasicus rises towards the end. Such relatively 'flat' calls resemble the evenly-pitched call of tristis and are quite different from the rising 'hweeet' of collybita and abietinus.
The calls of menzbieri in sonogram 'l' and the calls recorded at Kelardasht and Kiaser in Iran in April 2017 have a profile which is relatively straight initially but rises slightly and then falls towards the end, creating the semblance of a subtle 'terminal arch'. The slight rise and fall of these calls hints at a shallow 'sweeoo' sound (see footnote) but with the upward inflection well towards the end of the call i.e. an even tristis-like start progressing into a terminal 'sweeoo'.
The calls in the menzbieri sonogram provided by Helbig et al. are unequivocally menzbieri, based upon a recording at Kopet Dagh by Irina Marova, and these calls are a very close match with those recorded at Kelardasht and Kiaser during April 2017. More recently, Cedric Mroczko placed on xeno-canto (xc513431) recordings of chiffchaffs from Alagol, NE Iran, on the border with Turkmenistan. The breeding form here would be menzbieri but, as the recordings were made in November, there remains a possibility that some other taxon was involved. However, the sonogram (Figure 7) again shows a rising frequency towards the end and then a fall, the latter having a somewhat longer tail (descending to lower frequency) than in the Iranian recordings from April 2017 and Irina Marova's recordings from Kopet Dagh.
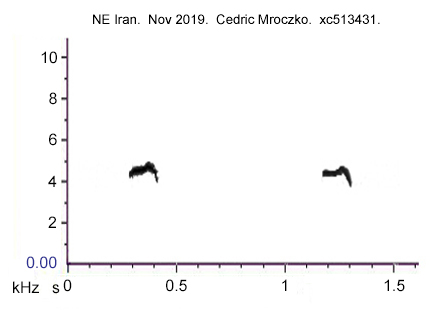
Figure 7.
Calls with an even more prominent 'terminal arch' and longer 'tail' have been recorded in winter in both Azerbaijan and Kuwait (thus wintering or potentially on passage and hence provenance unknown -see article on Chiffchaffs wintering in Kuwait). Such 'prominent terminal arch' calls have also been recorded in Turkey (and are thus significantly different from the descending frequency in brevirostris sonogram 'i' in Fig. 6 above). The frequency span of these 'terminal arch' calls recorded in Azerbaijan, Kuwait and Turkey is significantly greater than in the calls reliably attributed to menzbieri in Iran though calls with a more-comparable frequency span have also been recorded in Azerbaijan - but again in winter (see Chiffchaffs wintering in Kuwait). In summary, a call which begins on an even pitch but concludes with a shallow 'terminal arch' is a feature of Chiffchaffs in the breeding season and reliably assigned on range to menzbieri. However, 'terminal arch' calls with a more prominent arch and longer 'tail' have been recorded from Chiffchaffs in Kuwait, Azerbaijan and Turkey, though other types of calls have also been recorded from the last two. It is important to consider the season at which calls have been recorded, as in some instances birds on passage or wintering may be involved. It remains to be determined whether 'terminal arch' calls figure in the repertoire of all the southern taxa and, if so, whether the prominence of the 'terminal arch' is variable within a given taxon or whether it differs between the taxa.
In particular, reliable data on the call repertoire of caucasicus appears limited at present. Complications arise in respect of the calls attributed to caucasicus in the sonograms published by Helbig et al. (lower section of Figure 6 above). The first call in sonogram 'k' was recorded at Mount Elbrus in the Greater Caucasus by J. Martens, thus well inside the perceived range of caucasicus. This call has an upward inflection towards the end, recalling the opening part of a 'terminal arch' but (apparently) lacking the subsequent downturn and 'tail'. Careful inspection suggests just a hint of a short downward 'tick' at the end. It is conceivable that a downward tail has been truncated, lost or omitted from a 'terminal arch' call, as a result of individual call vagaries or simply become too faint to register owing to volume or recording circumstances. Alternatively, it may be an established call in the repertoire of caucasicus that has 'evolved' from a 'terminal arch' call. However, the attribution to caucasicus of the second and very straight call in Helbig et al's sonogram 'k' is questionable. It is.from a recording made by J. Martens in the Alamdeh region of northern Iran, which is in the same general region as Kelardasht, i.e. a region where the breeding form is now inferred to be menzbieri according to the genetic studies of Raković et al. Given that the sonogram designations are correct, then it appears that straight calls and calls with a terminal arch have both been recorded in a region of Iran where the breeding form is now deemed to be menzbieri. Correspondingly, it seems that only the first (and somewhat enigmatic) call in section 'k' of the sonograms presented by Helbig et al. is unequivocally from within the heartlands of caucasicus. This highlights questions over both the precise distribution of the two taxa and the extent of individual variation and overlap in calls within each taxon. Note also that the calls I recorded at Vorotan in Armenia in May 2018, a region where caucasicus and menzbieri converge according to Raković et al. (2019, also have quite an even pitch.
There are a few other published calls (and sonograms) attributed to caucasicus which show a 'terminal arch' but they are from the Lesser Caucasus, a region where caucasicus and menzbieri are now deemed to converge and overlap according to Raković et al, so the subspecies involved in these instances is ambiguous. Up to April 2024, apart from Marten's recording used in the first call in Helbig et al.'s sonogram 'k' (figure 6), I was unable to locate any recordings / sonograms of caucasicus calls from the Greater Caucasus. However, on April 19th 2024, Albert Lastukhin recorded calls attributed to caucasicus at Lechinkai in the Russian Federation and some 50km north of the border with Georgia. This location is within the Greater Caucasus and thus firmly within the range of caucasicus. His recordings can be found on xeno-canto as xc902028 and xc903809 and sonograms show that the calls are relatively straight. Following from this association between caucasicus and a straight call, the straight calls I recorded at Vorotan (a region where caucasicus and menzbieri converge according to Raković et al.) might be attributable to caucasicus (see Figure 8). However, the similarly straight calls recorded by Martens at Alamdeh in the Iranian Elborz (see second call in Helbig et al.'s sonogram 'k') are from well within the range now associated with menzbieri by Raković et al. (2019). Pro tem, the conclusion would appear to be that the call repertoires of both menzbieri and caucasicus may include 'straight' calls. These straight calls are particularly tristis-like, both in sound and in sonograms. Further recordings from the Greater Caucasus and across the newly inferred range of menzbieri are needed to elucidate typical calls and call variations within the Chiffchaffs breeding in these regions.

Figure 8.
In summary, valuable data on the vocalisations of the southern forms of Chiffchaff are emerging, including the significance of 'rising appendix' notes in songs and 'terminal arch' calls (neither of which is found in collybita or abietinus). There is a prima facie case that the standard call of menzbieri starts on an even pitch but concludes with a shallow 'terminal arch'. A 'flatter' or 'straighter' call may also figure in menzbieri's repertoire, while a recent recording from the Greater Caucasus in the Russian Federation (xc902028) indicates that a straight call is found in caucasicus and may be its typical call. However, the number of recordings from this heartland area of caucasicus remains very limited. 'Terminal arch' calls with a more-prominent arch (greater rise in frequency and longer 'tail') have been recorded in Turkey (breeding form brevirostris).
Calls with a prominent terminal arch have also been recorded in Kuwait (wintering birds) and Azerbaijan (where breeding birds potentially involve caucasicus or menzbieri but dates of recordings do not exclude migrants or wintering birds). Thus, the range of calls employed by menzbieri, caucasicus and brevirostris requires more investigation, including potential overlap and any significance in the frequency span ('prominence') of the 'terminal arch'. For examples and discussion of 'terminal arch' calls recorded recently in winter and /or on migration in the UAE and Saudi Arabia, see Motteau et al. (2022) and Babbington et al. (2024).
Longer series of recordings in the core breeding season are necessary, from Turkey, Georgia, Armenia, southern Russia, throughout Iran and from the Kopet Dagh area of Turkmenistan.
For further information on call variation and 'alternative' calls (including 'sweeoo' calls) among the northern subspecies see: <Alternative calls of Common Chiffchaff>.
Chiffchaff 'tret' calls
In an intriguing paper, Ivanistkii et al. (2020) have reported that the short, dry 'tret' calls, which Chiffchaffs interpose between the strophes in their songs, also differ between the various taxa of the chiffchaff complex (including here the 'Mountain Chiffchaffs' lorenzii and sindianus). The exception was that the 'tret' calls of collybita and abietinus were indistinguishable and there was a proviso over brevirostris, which (at least during their study) used such calls relatively infrequently. The authors suggest that the 'tret' calls of abietinus and caucasicus are sufficiently distinct to provide an aid to clarifying their geographical distributions. (Also fascinating is their finding that the 'tret' calls of tristis differ between central Siberia and NE Yakutia.)
Summary
The text below provides a summary of the the more detailed analysis above. It can be used as a 'refresher' for subsequent consultation but any 'third party' reference to this item should be based upon a thorough reading of the full documentation above.
Observations, photographs and sound-recordings of Common Chiffchaffs were made in April 2017 in the Elborz Mountains of Iran (east to Kiaser) and in May 2018 at Dilijan and the Vorotan Valley near Goris in Armenia. Based upon published distribution maps at the time, it was presumed that the Armenian birds would be caucasicus while the Iranian birds were left indeterminate between caucasicus or menzbieri. Armenia and N. Iran 'bridge' the ranges of currently designated caucasicus and menzbieri but their precise ranges remain undefined. Subsequently, genetic analyses conducted by Raković et al. (2019) detected a distinct clade around Transcaucasia, east and south of the brevirostris / caucasicus clade. Confirmatory data, including samples throughout N. Iran and Kopet Dagh, were still required for analysis but the authors inferred that this clade equated with menzbieri and that the range of this form included not only Kopet Dagh but the entire Hyrcanian forest of N. Iran and into southern Armenia as far as Goris. This implies that Chiffchaffs observed in N. Iran in April 2017 would be menzbieri while those at Dilijan in Armenia in 2018 were from the southern limits of caucasicus and those at Vorotan were in the region where the ranges of caucasicus and menzbieri converge and overlap. If indeed the range of menzbieri is so much more extensive than hitherto implied and extends so far west then, presumably, the prospects of extralimital occurrences in the Middle East and parts of Europe are correspondingly enhanced. [See Postscript below.]
It is hoped that the material gathered here may contribute to discussions of the characters, distribution and taxonomy of the southern subspecies of Common Chiffchaff. Common Chiffchaffs in Armenia and in the Elborz Mountains of Iran, at least as far east as Kiaser, share brown but grey-tinged upperparts and distinctly whitish underparts. Olive is also apparent on the upperparts but is restricted and comprises scattered streaking rather than an overall suffusion. The upperparts of the Armenian birds at Dilijan (perhaps caucasicus) appeared a little more olive than those of the Iranian birds (menzbieri following Raković et al. 2019) - especially considering the notionally fresher plumage of the Iranian birds, being in April, compared with May for the Armenian individuals. The decidedly whitish ground colour of the underparts is common to the Chiffchaffs in Armenia and Iran and was among the most distinctive features in the field. Yellow streaking is very limited and only detected at close range. The fringes to the remiges and rectrices, however, are distinctly yellowish-olive. The legs and feet are densely black. Such features have been associated with the form caucasicus (Loskot 1991), as discussed by Copete & López (2013), but are also compatible with comments on the appearance of menzbieri in the type description (Shestoperov 1937) and in discussions by Watson (1962), Marova & Leonovich (1997) and Loskot (2002). Potential morphological distinctions between caucasicus and menzbieri require further research.
A deficit of olive and yellow in the body plumage but combined with olive-yellow fringes to the flight feathers is no doubt the source of analogies which have been made between menzbieri and 'fulvescens', with which menzbieri was included historically by Vaurie (1954) and Williamson (1962), despite their distributions being separated by over 1500km. There is a suggestion of 'fulvescens' in the appearance of the of the upperparts of Iranian-designated menzbieri but the Iranian chiffchaffs have more conspicuous yellowish-olive fringes to the wing-feathers and tail-feathers, greyer-tinged upperparts and much whiter underparts (lacking the strong buff wash on the flanks of tristis s.l.).
Recordings of songs from the Elborz Mountains of Iran (east to Kiaser) in 2017 and from Armenia in 2018 are generally in keeping with sonograms attributed to menzbieri and caucasicus published by Helbig et al. (1996) and Clement et al. (1998). A note with a rising appendix, positioned wholly above the base frequency of the initial down-stroke and rising to between 40% and 60% of the note's overall peak frequency, is distinctive in sonograms of songs of Chiffchaffs from both regions (implicating menzbieri and caucasicus but not found, or rarely so, in the songs of other subspecies, including brevirostris). There are two forms of this style of note, one in which the rising appendix is evenly ascending ('straight'), the other in which it has a 'saw-tooth' structure. Both appear in the songs of menzbieri and caucasicus (Marova et al. 2021) but in my own recordings the 'saw-tooth' appendix was confined to the recordings from Iran. Intriguingly, using a multivariate analysis of song parameters, Marova et al. (2021) concluded that Caucasian menzbieri song clusters with that of caucasicus but differs from Kopet Dagh menzbieri.
To the ear, the calls of the Chiffchaffs in both Iran and Armenia are similar to the call of tristis. Nevertheless, most sonograms of calls from Chiffchaffs in Iran, within the wider range now attributed to menzbieri by the genetic clades identified by Raković et al. (2019), reveal an even pitch initially but leading into a shallow rise and fall at the end, creating a shallow 'terminal arch'. A more-prominent 'terminal arch' (with greater frequency span and longer downward 'tail') is apparent in recordings of one of two call-types attributed to brevirostris in Turkey and calls of some Chiffchaffs, of unknown subspecies currently, wintering in Kuwait and Azerbaijan (see <here>).
Calls I recorded in the Vorotan Valley in Armenia in 2018 (an area where caucasicus and menzbieri converge according to the genetic studies of Raković et al. 2019) show a relatively even pitch. Calls recorded by Albert Lastukhin in April 2024 in the Greater Caucasus, the core breeding range of caucasicus, also comprise relatively straight calls (xeno-canto: xc902028 & xc903809). Thus, a straight call seems to be associated with the breeding range of caucasicus and this may include some chiffchaffs in the region where caucasicus converges with menzbieri. However, a flatter call attributed to caucasicus in the sonograms published by Helbig et al. (1996) was recorded in the Alamdeh region of N Iran, a region where the breeding form of Chiffchaff is now equated with menzbieri by the genetic studies of Raković et al. (2019). Thus, both evenly pitched and (shallow) 'terminal arch' calls appear to have been recorded from an area of N Iran well within the region now associated with menzbieri.
In summary, there is a prima facie case that the standard call of menzbieri starts on an even pitch but concludes with a shallow 'terminal arch'. A call with a more prominent 'terminal arch' is found in one call associated with brevirostris. A 'flatter' or 'straighter' call is found in calls attributed to caucasicus. However, this situation is complicated by indications that the full repertoire of calls of the three forms may include more than one call type and a degree of overlap.
Further research across the full ranges of all three southern taxa is required, to determine their precise distributions, their morphological and vocal characteristics and how these may differ or overlap between the forms.
However, the geographical distribution of the southern forms as a group, the frequent 'whiteness' of their underparts, their tristis-like calls and the presence of distinctively-shaped notes in sonograms of their songs, set them aside from typical abietinus and collybita. The differences can be subtle and require further elucidation but clearly have significance. They are an intriguing element of the diversity of Phylloscopus collybita and further study of the intriguing Chiffchaffs found in the region is very much to be encouraged.
The form 'fulvescens' was described from migrants collected in Turkestan (Severtzov 1873) but the name and breeding range were equated by Sushkin and others with western Siberia. Later, Vaurie (1954) included in 'fulvescens' chiffchaffs from much further south, in SW Transcaspia and NE Iran. This was repeated by Williamson (1962), though he noted that this was a 'curiously disjunct distribution' and he also subsumed 'fulvescens' with tristis. Locations noted by Ticehurst (1938) of individuals with intermediate 'riphaeus' traits (deemed to be 'on passage', presumably because they were assumed to originate from the tristis / abietinus overlap zone) included the south coast of the Caspian and the Elborz Mountains in Iran. Chiffchaffs breeding in woodlands in NE Iran and Kopet Dagh in Turkmenistan are indeed greyer above, whiter below and deficient in yellow compared with typical abietinus but lack other features of tristis / 'fulvescens'. Their song is much like collybita / abietinus but their call is similar to tristis (see main text). They are now designated as menzbieri. At the time of Ticehurst's monograph (1938), menzbieri had only recently been described (Shestoperov 1937) and was not included (while caucasicus was not described (by Loskot) until 1991). As late as 1992, this confusion over menzbieri persisted, with BWP6 opting to include it within tristis but noting that 'it may belong to abietinus'.
The implication from Raković et al. (2019) of a much-enlarged breeding range for menzbieri raises the possibility that the taxon's wintering range might also be more extensive than hitherto supposed. In the UAE, Chiffchaffs with menzbieri DNA were trapped in March 2019 (one) and December 2020 (two) - Motteau et al.( 2022). As these comprised three of four Chiffchaffs genetically analyzed (the other being abietinus), the inference was that menzbieri is a regular component of the Chiffchaffs wintering in the UAE. Thus, the much-wider breeding and wintering ranges of menzbieri implied by the studies described above raise the prospect that menzbieri could reach Europe. Chiffchaffs with brevirostris / caucasicus mtDNA have already reached The Netherlands and Germany (van der Spek & de Knijff 2021). In Saudi Arabia, Chiffchaffs bearing brevirostris / caucasicus mtDNA have been detected (Babbington et al. 2024, where useful photographs and sonograms can also be found). As the calls of southern forms are very similar to the call of tristis (nearly identical in cases), this emphasises that identification of extralimital Chiffchaff subspecies should always embrace a full suite of features : morphology and vocalisations.
Acknowledgments
I would like to thank Ali Alieslam, Steve Rooke and Rob Tizard for discussions in the field and provision of their excellent photographs of Chiffchaffs in Iran; José Luis Copete and Lars Svensson for comments (in litt.); Irina Marova for comments (in litt.) and for providing a copy of her paper with V. V. Leonovich on menzbieri at Kopet Dagh; Timo Janhonen for providing information on Chiffchaffs he observed and sound-recorded in Azerbaijan in March 2012; Oscar Campbell and Jem Babbington for discussions of the appearance and vocalisations of Chiffchaffs wintering in the UAE and Saudi Arabia, respectively; Antero Lindholm for discussions of Chiffchaffs in Georgia and surrounding regions; Marko Raković and his co-authors for providing 'open access' to their paper via the PLoS publishing organisation; Martin Collinson for clarifying certain genetic issues in that paper; Daniel Duff for more-general discussion of the potential traits of southern forms of Chiffchaff; Roger Riddington for permission to re-use sonograms published in British Birds.
References
Babbington, J., Roberts, P., Senfeld, T. & Collinson, J. M. 2024. Subspecies of Common Chiffchaff in Saudi Arabia. Brit Birds 117: 438 - 455.
Clement, P., Helbig A. J. & Small, B. 1998. Taxonomy and identification of chiffchaffs in the Western Palearctic. Brit. Birds 91: 361-376.
Clements, J. F., Schulenberg, T. S., Iliff, M. J., Roberson, D., Fredericks, T. A., Sullivan, B. L. & Wood. C. L. 2018. The eBird/Clements checklist of birds of the world: v2018. Downloaded from http://www.birds.cornell.edu/clementschecklist/download/
Copete, J. L. & López, F. 2013. Identificación de subespecies en el mosquitero común, in Rodríguez, N., García, J. & Copete, J. L.(eds). El mosquitero iberico. Leon.
Cramp, S. (ed.) 1992. The Birds of the Western Palearctic. Vol. 6. Oxford.
Dean, A. R. Updated 2021. Chiffchaffs wintering in Kuwait : which subspecies? https://deanar.org.uk/general/articles/KuwaitChiffchaffs.htm
Dementiev, G. P. 1948. Material on ornithology of North Iran. Trudy Byuro kol'tseveniya 7: 177 - 194. [In Rusian].
Gill, F., Donsker, D. & Rasmussen, P. (Eds). 2020. IOC World Bird List (v10.1). doi : 10.14344/IOC.ML.10.1.
Grishchenko, A. V., Tsvelykh, A. N. & Yablonovska-Grishchenko, E. D. 2016. Song repertoire and origins of Crimean population of Chiffchaff, Phylloscopus collybita (Sylviidae). Vestnik zoologii 50: 89-92.
Helbig, A. J., Martens, J., Seibold, I., Henning, F., Schottler, B. & Wink, M. 1996. Phylogeny and species limits in the Palearctic Chiffchaff Phylloscopus collybita complex: mitochondrial genetic differentiation and bioacoustic evidence. Ibis 138: 650-666.
Ivanitskii, V., Ilina, I. & Marova, I. 2020. The sounds between the strophes: different chiffchaff taxa perform different tret calls in their song. Bioacoustics doi: 10.1080/09524622.2020.1796788.
Loskot, V. M. 1991. A new Chiffchaff subspecies (Aves, Sylviidae) from the Caucasus. Vestnik zoologii, Kiev 3: 76 -77. [In Russian]
Loskot,V. M. 2002. On the type specimens of Sylvia curruca caucasica Ognev & Bankowski and Phylloscopus collybita menzbieri Shestoperov (Aves: Sylviidae). Zoosystematica Rossica 10: 413-418.
Marova, I. M. & Leonovich, V. V. 1997. Mysterious Chiffchaff from the Kopet Dagh: ecology, vocalization and relations of Phylloscopus collybita menzbieri. Zoologichesky journal 76: 735-742. [In Russian with English summary]
Marova, I., Ilyina, I., Kvartalnov, P., Grabovsky, V., Belokon, M., Solovyova, E. & Ivanitskii, V. (2021). From the Bosporus to Kopet Dagh: Morphological, Genetic and Bioacoustic Variation in the Chiffchaff in Turkey, the Caucasus and Western Turkmenistan. Ardea 109. 10.5253/arde.v109i3.a3.
Motteau,V., Campbell, O., Senfeld, T., Shannon, T. J., Collinson, J. M. & Lloyd, S. 2022. First occurrence of menzbieri Common Chiffchaff in the United Arab Emirates. Brit. Birds 115: 500 - 509.
Raković, M., Neto, J. M., Lopes, R. J., Koblik, E. A., Fadeev, I. V., Lohman, Y. V., Aghayan, S. A., Boano, G., Pavia, M., Perlman, Y., Kiat, Y., Ben Dov, A., Collinson, J. M., Voelker, G., & Drovetski, S. V. 2019. Geographic patterns of mtDNA and Z-linked sequence variation in the Common Chiffchaff and the 'chiffchaff complex'. PLoS ONE 14(1): e0210268. (https://doi.org/10.1371/journal.pone.0210268)
Shestoperov, E. L. 1937. Birds (Aves). Opredelitel' pozvoniJchnykh zhivotnykh Turkmenskoi SSR [Keys to Vertebrata of the Turkmenian SSR], 4: 331 p. Ashkhabad - Baku. [In Russian]
Shirihai, H. & Svensson, L. 2018. Handbook of Western Palearctic Birds : Passerines, volume 1. London.
Svensson, L. 1992. Identification Guide to European Passerines. Fourth edn. Stockholm.
van der Spek, V. & de Knijff, P. 2021. Migrating Chiffchaff taxa in The Netherlands: a ten-year genetic study. Dutch Birding 43: 120 - 131.
Vaurie, C. 1959. The Birds of the Palearctic Fauna. Order Passeriformes. London.
Watson, G. E. 1962. A re-evaluation and redescription of a difficult Asia Minor Phylloscopus. Ibis 104: 347 - 352.
xeno-canto. (Recordings of Chiffchaff taxa.) https://xeno-canto.org/
Dean, A. R. 2017 (Updates to 2024). Southern forms of Chiffchaff Phylloscopus collybita : observations from Iran and Armenia. https://deanar.org.uk/general/articles/IranChiffchaffs.htm
|
|