
|
Site home-page |
'Siberian Chiffchaff' Phylloscopus collybita tristis: discussion and photo gallery. |
Parts of this item are based upon:
Dean, A. R. & Svensson, L. 2005. 'Siberian Chiffchaff' revisited.
British Birds 98:
396-410
&
Dean, A., Bradshaw, C., Martin, J., Stoddart, A. & Walbridge, G.
2010. The status in Britain of
'Siberian Chiffchaff'.
British
Birds 103: 320-338.
References to genetics are unavoidable but, in this text, they are from a non-geneticist's perspective and subject to correction.

Plate
1.
Two 'classic' Siberian Chiffchaffs Phylloscopus (collybita) tristis
Classic tristis are relatively 'colourless' or 'monotone'
rather than truly pale (as is often inferred). They lack yellow away from the
bend of the wing and lack olive in the crown and mantle.
Additionally, they have
a characteristic tan-brown hue to the upperparts and a rich buff suffusion to supercilium, ear-coverts, sides of breast and flanks,
evident here in both a spring and an autumn individual.
The Common Chiffchaff Phylloscopus collybita has a broad distribution across Europe and east to Siberia, with six subspecies currently recognised by most authorities. Broadly speaking, the nominate form collybita of western Europe is replaced by abietinus in eastern Europe and by tristis in Siberia. Further south three additional forms have been described : brevirostris in western and northern Turkey, caucasicus in the Greater and Lesser Caucasus and menzbieri, historically considered confined to Turkmenistan and north-eastern Iran. The form tristis is colloquially known as 'Siberian Chiffchaff' and is treated by some authorities as a distinct species. Its breeding range extends from the Mizen Basin (west of the Urals) east across Siberia to the Kolyma River. The taxonomy and diagnosis of ‘Siberian Chiffchaff’ have long been beset by confusion and divided opinion, with considerable uncertainty surrounding both its systematic and morphological limits. Investigations based upon mitochondrial DNA (mtDNA) confirmed 'genetic distance' between the currently designated forms within the chiffchaff complex and helped to clarify species boundaries. However, whether tristis warranted recognition as a distinct species remained inconclusive (Helbig et al. 1996; Clement, Helbig and Small 1998). The origins of variation in the appearance of Chiffchaffs in the west of the range of tristis have also remained contentious, attributed variously to hybridization between tristis and abietinus or to individual, clinal and even taxon-level variation. Original genetic studies which were confined to mtDNA, which is inherited solely via the female line, led to differing interpretations. Marova et al. (2009, 2013) concluded that hybridization was widespread in the region of sympatry, while studies of migrants in the Netherlands by de Knijff et al. (2012) led to counter suggestions that the plumage of tristis was naturally more-varied than had been implied hitherto. The extent to which vocalisations are diagnostic has also been obscured by the issues of call variation within all taxa, hybridization with abietinus and the very tristis-like calls given by forms of Chiffchaff found in the Middle East and the Caucasus. More recently, techniques for examining 'whole genome sequence data' have been developed and applied to Chiffchaffs in the zone of sympatry between tristis and abietinus in the Urals and Arkhangelsk region (Shipilina et al. 2017). Significant genetic mixing between tristis and abietinus has been confirmed, with Shipilina et al. concluding that this was 'the driving force underlying trait variation' and was also implicated in vocal anomalies. The following pages explore these issues and their implications for field identification of 'Siberian Chiffchaff'.
Topics
Appendix 1. The 'type descriptions' of tristis and 'fulvescens'
Appendix 4. Historical records of 'Northern' or 'Eastern' Chiffchaffs
Appendix 5. Colour-corrected images of tristis from Dean & Svensson (2005)
Photos
Case studies
Any text discussing the appearance of Chiffchaffs is immediately beset by the issue of colour nomenclature. Several colours and shades - olive, yellow, brown, buff, grey and white - are exhibited by most Chiffchaffs to one degree or another. All subspecies of Chiffchaff vary in appearance to a degree but it is the extent and intensity of these key hues and the degree to which they are admixed that lead to a characteristic appearance for each subspecies and define the appearance of what might be termed 'classic' individuals. Hence, colour nuances are central to discussions of plumage traits in Chiffchaffs. However, the absence of a universally recognised and accepted 'colour nomenclature' adds yet further complexity to discussions of plumage colours and differences. Indeed, different observers often apply contradictory colour names to the same individual Chiffchaff while, conversely, identical colour names are often applied to two or more Chiffchaffs of quite different appearance. When discussing Chiffchaffs, it is near-impossible to avoid using the terms 'grey' and 'buff' but both of these suffer from especially diverse interpretations between different observers and this can and does lead to ambiguity and confusion.
In my own texts, I employ shorthand terms for certain colour liveries, such as 'olive and yellow', 'brown and buff' and 'grey and white'. Clearly, there is nothing very 'nuanced' about these terms and they are no more than labels of convenience. They are relative, not absolute terms, designed to indicate the key colours and shades which distinguish the Chiffchaffs to which they are applied. In a wider context, 'grey' can span any shade between white and black, as in the 'Kodak Grey Scale' below. In the term 'grey and white' livery, the label 'grey' is intended to convey a shade resembling 'pale neutral grey' in Smithe's 'Naturalist's Color Guide' or shades in the range 2 to 4 on the Kodak Grey Scale. 'Buff' is here used to designate the hue which infuses the breast and flanks of a truly 'classic' Siberian Chiffchaff (see plate 1). In this usage it lacks yellow and has a slightly 'rusty' tinge (in more detailed discussions in the text, the term 'rusty-buff' is sometimes employed). Thus, it signifies a somewhat warmer shade than is sometimes inferred by the term 'buff'. It essential that all these shorthand terms are interpreted with reference to the definitive and carefully chosen photographs from which they derive their meaning (see Plate 2 below). See also: Colour issues, 'moult & wear' and photographs .

Plate 2. A typical 'olive and yellow' collybita on the left, a typical 'brown and buff' tristis on the right and a 'grey and white' Chiffchaff in the centre (taxon much debated - see Casestudy1). Classic 'brown-and-buff' tristis lack olive and yellow in the body plumage (apart from yellow on the underwing coverts) while classic 'grey-and-white' Chiffchaffs lack the distinctive tan-brown and rusty-buff hues characteristic of classic tristis (see main text for further discussion and nuances). Chiffchaff plumages as a whole encompass a somewhat 'sliding scale' but terms such as 'brown-and-buff' and 'grey-and-white' are convenient 'shorthand' and encapsulate the relative 'colour balance' differences of archetypal forms. The image above provides a reference for the application of these terms in the texts on this site. A 'pixel map' from the mantle of each of the three Chiffchaff images above can be seen in Colour issues and photographs.
Chiffchaff plumages vary individually and a diminution of olive and yellow from west to east is frequently cited, though to what extent this is truly clinal warrants further investigation. A complete lack of yellow away from the underwing has been a defining characteristic of tristis since the type description by Blyth in 1843 (which can be examined in Appendix 1). Witherby et al. (1938), in 'The Handbook of British Birds', and Ticehurst (1938), in 'A Systematic Review of the Genus Phylloscopus', also emphasised the restricted yellow in the plumage while also noting the browner upperparts compared with abietinus. More recently, the most-frequently-cited reference to the definitive features of tristis has been the description provided in Lars Svensson's 'Identification Guide to European Passerines' (1992, 2023), often dubbed the 'Svensson criteria', which may be summarised as:
supercilium, eyelids and cheeks buffish (no yellow)
no yellow on underparts apart from axillaries and bend of wing
cheeks, sides of throat and breast tinged rufous-buff
no green on mantle, nape or crown
rump, edges of wing-coverts, remiges and rectrices tinged olive-green
black or blackish tarsi
small dark bill
Additionally and often under-emphasised in the past is:
a distinctive tan-brown tinge to the upperparts
To these morphological criteria might be added:
unequivocal delivery of the classic evenly-pitched, monosyllabic and plaintive call ('eeep' or 'iiihp')
While the deficiency of yellow and olive is fundamental, the brown component in the upperparts is also characteristic, often with a small but influential russet component, at its most distinctive producing a 'tan' or 'khaki' tinge (see HSB analyses). Equally, the buff hue to the supercilium, eye-lids and cheeks generally extends to the sides of the breast and flanks, with only the central belly and undertail coverts being rather whiter (see plate 1). Thus, the plumage as a whole has characteristic 'brown and buff ' hues. In the field, evaluation of precise hues requires that observations are extensive, are conducted at close range and in suitably neutral light conditions. In brief views and adversely bright conditions, the true hues can be diluted and lead to a misleading impression. When examined in appropriate conditions, which reveal the colours accurately, any individual which lacks these brown and buff hues entirely is unlikely to be tristis. The plumage of 'classic' tristis recalls that of Mountain Chiffchaff P. lorenzii, though the latter has a darker crown and a whiter and more striking supercilium, which frequently bridges the forehead (it also noted in some texts that lorenzii has more subdued olive fringes to the flight feathers than tristis but such olive fringes are a rather variable feature even in tristis, and tend to more obvious on individuals from west of the Yenisey).
The 'Svensson criteria' were intended to define 'classic' individuals and hence offer high 'confidence limits' in any diagnosis. However, only east of the Yenisey do all individuals lack olive on crown and mantle and exhibit yellow only on the bend of the wing and underwing. West of the Yenisey, while some individuals match the criteria for 'classic' individuals, others have very slight traces of 'misplaced' yellow and olive. Such individuals have become known as 'fulvescens'.
West of the Yenisey, the appearance of Chiffchaffs is less consistent. As noted by Ticehurst (1938), Russian authors including Sushkin (1925) designated Chiffchaffs from the region west of the Yenisey as fulvescens, primarily on the basis of brighter olive on the rump and the fringes to the remiges compared with classic tristis. Ticehurst suggested that individuals with such olive enhancements were to be found throughout the range (not just the west) and consequently questioned the validity of 'fulvescens'. It is worth noting, however, that 'classic' tristis frequently exhibit very little if any olive on the fringes to the flight feathers and hence lack both yellow and olive almost entirely (see Plate 1 for both a spring and an autumn individual exhibiting truly classic appearance). From examination of specimens, Svensson (1992) noted that some individuals in the region west of the Yenisey match the ‘classic’ birds from farther east (as embraced by the traditional 'Svensson criteria') but others have traces of yellow on the underparts, the supercilium and often on the upper part of the eye-ring, and traces of olive in the crown and mantle. In recent literature, as well as brighter olive hues to the wing-edges and rump, the features associated with the term 'fulvescens' have come to include 'additional' or 'misplaced' olive and yellow in comparison with classic tristis, with yellow in the supercilium and upper eyering perhaps the most frequent manifestation of 'fulvescens' traits in this scenario. Candidate Siberian Chiffchaffs reaching western Europe not infrequently display such 'fulvescens' traits (see plates 3 & 4).
The name 'fulvescens' derives from the type description of Severtzov (1873), thus well before Sushkin's discussion of Chiffchaffs from the region west of the Yenisey, and was based on a series of migrants collected in Turkestan. Thus, they were not obtained from the West Siberian Plain and, although Severtzov suggested their breeding grounds might be in western Siberia, their natal origins were uncertain. See Appendix 1 for a transcription of the type description (kindly provided and translated by the late Dr Vladimir Loskot, past Curator of the Ornithological Department, Zoological Institute, Russian Academy of Sciences, St. Petersburg). There are potential ambiguities arising from linguistics and colour nomenclature e.g. the English translation of the type description results in the phrase 'rusty-yellow' for the supercilium and underparts, suggesting the presence of 'additional' yellow compared with 'classic' tristis. However, as this term is applied to the ground-colour (not an additional tinge or streaking), 'rusty yellow' in translation may well refer to the 'buff' hues found on the supercilium and underparts in all tristis-types. This likelihood is enhanced by the use of 'sulphur yellow' to describe 'unclear streaking' on the underparts of some juveniles (and also as the colour of the underwing coverts). This streaking on the underparts is described as 'greyish' in other juveniles, to which the name 'naevia' was applied. Despite these historical uncertainties, the term 'fulvescens' has now become equated with Chiffchaffs differing from 'classic' tristis in the presence of traces of 'additional' olive and yellow, while their designated origin is the region between the Urals and the Yenisey. As specimens from the now-designated breeding range of 'fulvescens' (primarily the West Siberian Plain) are limited (Shirihai & Svensson 2018), a more-comprehensive determination of the features and the variability of chiffchaffs from the region between the Urals and the Yenisey is still required, including just how far east traces of 'additional' yellow are encountered. See also Appendix 2 for a discussion of the geographical distribution of 'fulvescens' as treated in some earlier texts and the confusion with chiffchaffs now designated as menzbieri.

© A. Warr
Plate 3. Siberian Chiffchaff with 'fulvescens' traits, Powick, Worcs, February 2015
The 'Siberian Chiffchaff' in Plate 3 shows the 'tan-brown' upperparts, buff suffusion on underparts and warmer buff ear-coverts matching a 'classic' tristis but the yellow tinge
to the upper eye-ring, prominent yellowish-olive fringes to remiges and perhaps the extent of the pale base to the bill are all 'fulvescens' traits as the term is now interpreted.
Compare this individual carefully with those in Plate 1.
Full details of the Powick indivdual, and a more 'classic' individual also present at the site, can be found on Andrew Warr's blog <here>.

Plate 4. Two Siberian Chiffchaffs, left-hand individual displaying 'fulvescens' traits
Both individuals in Plate 4 show the characteristic
'brown and buff ' hues of Siberian Chiffchaff but, in the Powick individual, note the presence of
distinct yellow tinge in upper eye-ring and evident
yellowish-olive fringes in the visible 'shoulder' area.
Also the pale base to the bill, though this is not exclusive to 'fulvescens'.
The origins and implications of 'fulvescens' traits have been the subject of debate for many years. A long-standing opinion is that traces of 'additional' or 'misplaced' olive and yellow arise from introgression from abietinus, as the ranges of the two forms are contiguous and, indeed, overlap in an area west and north from the Urals. The potential for widespread hybridization, with a significant number of F1 hybrids and early backcrosses, lies only in this region of overlap i.e. the sympatric region. A low level of introgression may extend east across the allopatric West Siberian Plain but the levels of genetic mixing in the two regions are not comparable and the appropriate taxonomic treatments may also differ. Thus, to facilitate discussion, the name 'fulvescens' should be restricted to individuals matching the appearance of chiffchaffs from the allopatric region between the Urals and the Yenisey, which match classic tristis closely apart from slight 'additional' olive and / or yellow, often detectable only in the hand or in the highest-fidelity digital images. F1 hybrids and early backcrosses from the sympatric region around the Urals, frequently (but not inevitably) displaying more evidently intermediate morphology, are better referred to as 'riphaeus'. Ticehurst (1938) appreciated the distinction and used the two names in this sense colloquially, while noting that he did not advocate formal names for individuals which he regarded as variants of tristis ('fulvescens') and hybrids between tristis and abietinus ('riphaeus'), respectively. Clearly, F1 hybrids and early backcrosses ('riphaeus') cannot be designated as 'Siberian Chiffchaffs'. Unfortunately, discussions of introgression, hybridization and character variation across the range have been hampered by a widespread tendency to apply the name 'fulvescens' indiscriminately to all tristis-like Chiffchaffs differing from the 'classic' profile, whether from the region of sympatry around the Urals or from the allopatric West Siberian Plain. In Dean & Svensson (2005), Lars Svensson suggested that, pragmatically, 'fulvescens' might be better accommodated within tristis. This has been incorrectly interpreted by some subsequent authors. It should be noted that 'fulvescens' was used sensu stricto i.e. to refer explicitly to individuals found in allopatry in the West Siberian Plain, between the Urals and the Yenisey, where the level of introgression is low. The term 'riphaeus' was employed for individuals in the hybrid zone around the Urals, where genetic mixing was liable to be significant (i.e. 'riphaeus' equates with F1 hybrids or very early backcrosses). There was no suggestion that 'riphaeus' types could be accommodated within tristis.
Of course, this distinction applies at the populations level, and the level of genetic mixing in a given individual will vary. While the level of introgression of abietinus genes into the tristis population will be relatively low in the allopatric region and relatively high in the sympatric region, on-going backcrossing will presumably result in some individuals, even in the sympatric region, having a 'fulvescens'-like genetic composition i.e. a majority tristis-based genome. Thus, a 'fulvecens' type does not inevitably indicate an origin outside the sympatric zone. The issue of selective backcrossing will have a bearing on this (i.e. any 'mate selection preference' that F1 hybrids might exhibit, such as a preference to mate with tristis).
See genetic studies below.
Past descriptions of candidate Siberian Chiffchaffs frequently described them as being 'very pale and grey'. While tristis can appear rather 'monotone' or 'colourless' to observers for whom nominate collybita (with its prominent olive and yellow hues) is the norm, in general they are not especially pale when viewed in normal light conditions (see Plate 1), except for the whiter central belly and under tail coverts. However, when viewed in adversely bright light, they can be prone to 'colour dilution' (though this illusion is not restricted to tristis of course). Historically, it was very easy for such fleeting impressions to influence the public perception of tristis and to create a 'prior image' or 'expectation'. This could lead to instant judgements and override the essential more-prolonged evaluation of appearance in balanced light conditions. Dean and Svensson (2005) discussed in detail the appearance of classic tristis and 'fulvescens', noting that tristis approached in appearance Mountain Chiffchaff P. (sindianus) lorenzii and emphasising that 'fulvescens' differed from classic tristis principally in exhibiting slight 'additional' olive and yellow. Thus, descriptions of tristis-candidates which emphasised a 'distinctly pale and grey' appearance, with little mention of the characteristic tan-brown and buff hues, lacked key characteristics of the established appearance of both classic tristis and 'fulvescens' (as in Plates 1, 3 & 4 above). Increased and more-objective attention to the features of Siberian Chiffchaff followed this publication and revealed that, under adequate scrutiny, most tristis-candidates were, in fact, typical 'tan-brown and buff ' individuals, though 'fulvescens' traits were not uncommon. The high incidence of extremely pale tristis-candidates, lacking underlying tan-brown and buff hues, as inferred hitherto, had been exaggerated (Dean et al. 2010). The illusion of extreme 'pallor' resulted not only from fleeting impressions in the field (so-called 'morphing') but, not infrequently, this was exacerbated by too much reliance on digital photographs, which are far more susceptible to colour artefacts and colour dilution than has at times been acknowledged. Suitably extensive observations, conducted in more-neutral light conditions, confirmed that this overly pallid impression was transitory and that the more-enduring appearance in neutral light was quite in keeping with either classic tristis or 'fulvescens'.
With this more-balanced appreciation of the defining features of tristis becoming established, a further and unfortunate consequence was that some third-parties went on to claim that 'grey and white' Chiffchaffs (lacking evident 'brown and buff' hues) did not exist at all, within any subspecies, and were invariably an optical illusion. This was a non-sequitor. That tristis can appear deceptively pale and 'grey and white' in adverse light does not mean that Chiffchaffs with genuinely enhanced grey and white aspects to their plumage do not exist. The dismissal by some of Chiffchaffs with such 'grey and white' liveries has hampered discussions of plumage limits in both tristis and abietinus. Careful and prolonged observations have confirmed that, while infrequent, a small number of Chiffchaffs occurring in late autumn have an underlying appearance that is reasonably summarised as relatively 'grey and white'. Such 'grey and white' individuals exhibit reduced olive and yellow in their body plumage compared with typical abietinus and collybita, while there are often relatively bright and contrasting yellowish-olive fringes to the remiges and rectrices. At their most distinctive, they resemble one of the Bonelli's Warblers (Western P. bonelli or Eastern P. orientalis) whereas true tristis resembles Mountain Chiffchaff P. lorenzii. See centre bird in plate 2 and 'A case study from Scilly' for an example of a 'grey and white' style Chiffchaff. However, such 'grey-and-white' Chiffchaffs are not necessarily tristis. Dean & Svensson (2005) had already suggested that 'grey and white' Chiffchaffs may have more than one origin, viz.
paler extremes of tristis / 'fulvescens'
paler examples of abietinus from the eastern part of its range
intergrades between tristis and abietinus
variants among all subspecies of Chiffchaff
a taxon from an infrequently visited part of Chiffchaff's overall range which exhibits relatively 'grey and white' appearance (and also utters a tristis-like call)
Subsequently, the last of these options has been shown to be undoubtedly relevant, as certain southern forms of Chiffchaff, caucasicus and menzbieri, exhibit precisely these characters (see https://deanar.org.uk/general/articles/IranChiffchaffs.htm).
Three examples of such 'pale and grey' Chiffchaffs observed in Britain are shown in Plate 5 : one was trapped and photographed in the hand by Gary Crittenden in Highland in November 2008; the second I photographed on St Agnes, Scilly, in October 2011; the third was photographed by Martyn Pitt at Belvide Reservoir in Staffs, in December 2012. While there are subtle differences between them, all three exhibit fundamentally the same livery. On October 28th 2011, the St Agnes individual gave a rising abietinus/collybita-like call (pers obs) and also an inflected 'sweeoo', while the Staffs individual gave a 'quiet collybita style call throughout its stay' from December 1st - 3rd 2012 (per Steve Nuttall).

Plate 5. 'Grey and white' Chiffchaffs P. collybita.
As noted above, in their overall appearance such 'grey and white' Chiffchaffs are reminiscent of Western P. bonelli or Eastern Bonelli’s Warbler P. orientalis, though lacking such key Bonelli's characters as face-pattern, broken eyering, yellowish rump and bare parts. In Plate 6, the 'grey and white' Chiffchaff on St Agnes, Scilly, in October 2011 is compared with an Eastern Bonelli's Warbler in Turkey, photographed in the hand by José Luis Copete.

Plate 6. 'Grey and white' Chiffchaff P. collybita compared with Eastern Bonelli's Warbler P. orientalis
In plates 7 & 8, three different images of the St Agnes Chiffchaff are shown above images of Chiffchaffs photographed within the range of abietinus: Sweden (© Lars Svensson) and Finland (© Petteri Lehikoinen), respectively. The image from Sweden is reproduced from plate 240 in 'Siberian Chiffchaff revisited' (Dean & Svensson, 2005). The image from Finland is reproduced, with permission, from plate 4 in Copete. J. L. & López, F. 2013. 'Identificatiön de subespecies en el Mosquitero Comün (Phylloscopus collybita collybita, P. c. abietinus, P. c. tristis, P. c. brevirostris, P. c. caucasicus y P. c. menzbieri)'. The Scandinavian individuals were equated with abietinus by very experienced ringers for whom abietinus is the norm, with Petteri Lehikoinen commenting re the Finnish bird that this was supported by biometrics and call as well as location.

Plate 7. Three images of 'grey and white' Chiffchaff P. collybita, Scilly, Oct 2011. Compare with examples of 'grey' Scandinavian Chiffchaffs P. c. abietinus in Plate 8.

Plate 8. Two examples of relatively 'grey' Scandinavian Chiffchaffs P. c. abietinus. Compare with 'grey and white' Chiffchaff, Scilly, Oct 2011, in Plate 7.
Two images from Estonia below, by Uku Paal, are somewhat darker and not 'Bonelli's-like' but illustrate further the deficiency of olive and yellow exhibited by some autumn Chiffchaffs.

Plate 9. Two Chiffchaffs from Estonia, showing limited olive and yellow.
Individuals such as those in plates 5 and 8 have been observed and photographed carefully and their appearance confirmed as enduring and not transitory. Claims that 'grey and white' Chiffchaffs are invariably 'typical' tristis morphed by adverse light conditions are unsustainable. Further photos of the Scilly 'grey and white' Chiffchaff, together with images of a very 'brown and buff' tristis present at the same time, can be found in the case-study <here>, where the issue of so-called 'plumage morphing' is further addressed.
Debate about the appearance of abietinus thus became as contentious as that relating to tristis. Ringing studies in Britain (Collinson et al. 2018) and the Netherlands (van der Spek & de Knijff 2021) found that genetically confirmed abietinus and collybita in their samples were indistinguishable, including via biometrics, leading to the comment that, were it not for genetic differences, the validity of abietinus as a subspecies would be questionable. However, this conclusion was based upon samples trapped in limited geographical regions of western Europe. These and other issues are explored further within the following examination of genetic studies, which in many respects have assumed a central role in discussions of the chiffchaff complex.
In a 1500km long corridor extending from the southern Urals to the Archangelsk region, plumages are regularly encountered which include traces of 'misplaced' yellow in comparison with 'classic' tristis. They may be more
intermediate between classic tristis and abietinus than in the original application of the name 'fulvescens' sensu Sushkin (1925), as described above. Indeed, such more-evidently intermediate-looking individuals were designated as riphaeus (not 'fulvescens') by the Russian ornithologists who first studied them and some Russian authors (e.g. Buturlin & Dementiev 1937) described 'riphaeus' from the Urals region as a product of hybridization between fulvescens and abietinus. Marova et al. (1993, 2009, 2013) employed a combination of morphological, vocal and mtDNA data to study Chiffchaffs in the region of sympatry between tristis and abietinus from Arkhangelsk to the southern Urals mountains. The authors concluded that hybridization was widespread in the region of sympatry. However, they reported an imbalance in gene flow, with 88.5% of the individuals they deemed to be hybrids carrying the mtDNA haplotype of tristis. (Unfortunately, they applied the name 'fulvescens' to these hybrids in the sympatric region around the Urals, rather than the preferable 'riphaeus'.)In 2012, de Knijff et al. (2012) published an mtDNA-based study of Chiffchaffs trapped in the Netherlands. Ringers had been asked to assign to either abietinus or tristis trapped individuals deemed to differ in appearance from nominate collybita, based upon their appearance in the field. Subsequent analysis of mtDNA found that all individuals assigned by their ringers to abietinus returned tristis mtDNA. No individuals at all were found with abietinus mtDNA (though, as samples for genetic analysis had been selected on the basis of an appearance not suggestive of nominate collybita, some abietinus - which can be very like collybita - may have been missed). Thus, as birds assigned by their ringers to abietinus proved to have tristis mtDNA, this study concluded that some Chiffchaffs with tristis mtDNA exhibited a greater distribution of yellow and olive than was compatible with the traditional criteria for tristis. However, photos published with this study bring into question whether the characters for diagnosing tristis candidates were applied adequately by some of the ringers. For example, the individuals in plates 531and 533 are quite similar and, on the basis of the photos, they both meet the defining criteria for tristis morphotypes as listed above. Yet, the first was assigned as abietinus and the latter as tristis. Also, perhaps influenced by a paper on Siberian Chiffchaffs published by van den Berg (2009), which argued that hybridization between tristis and abietinus was rare 'if it occurred at all', this first of two Netherlands studies did not consider hybrids and 'fulvescens' and led to suggestions that the plumage of tristis was naturally more varied than had been implied hitherto.
In a second Netherlands study, van der Spek & de Knijff (2021) found that ringers' success in allocating Chiffchaffs to tristis in the field had improved significantly since the earlier study. All but one individual with tristis mtDNA were diagnosed as tristis in the field and the exception was a tristis morphotype which gave collybita/abietinus calls and hence was left unassigned. They also reported that further 'whole genome' studies would be conducted, to determine whether introgression was responsible for the presence of 'additional' olive and yellow in individuals assigned to abietinus in the earlier study but proving to have tristis mtDNA. However, a proviso here derives from the fact that there were very different levels of 'success' between the first and subsequent studies, in diagnosing tristis in the field. For further genetic studies, it is essential that the individuals in the earlier Dutch study attributed by their ringers to abietinus were indeed distinct in appearance from tristis and did exhibit 'mismatching morphotype and mtDNA' . As noted, on the basis of the published images, this is questionable in several instances from the earlier study. Genetic studies confined to the second batch of Chiffchaffs should be more well-founded.
Since mtDNA cannot identify hybrids directly (being inherited only from the maternal side), firmer resolution of the question of hybridization depended upon the development of techniques for examining the nuclear DNA of the forms. This proved elusive for several years but, in 2017, Shipilina et al. published a much-needed study, extending the earlier studies of Marova et al. to include 'whole genome sequence data'. This study confirmed that chiffchaffs within the sympatric region have 'a mix of genetic ancestry indicating extensive ongoing and past gene flow'. Additionally, mismatches were found between phenotype ('appearance') and haplotype (mtDNA) in the sympatric region but not among individuals from allopatric regions around the Yenisey. Contrary to earlier suggestions, Shipilina et al. concluded that natural plumage variation among thoroughbred tristis is limited, that ‘genetic admixture is the driving force underlying trait variation in the sympatric region’ and that intermediate-looking individuals are hybrids. In a comment included in Dean (2018), Daria Shipilina wrote: 'So far we have captured birds in three different (allopatric) locations in Siberia: Middle Yenisey, Lower Yenisey and Yakutia and didn't find any variation in phenotypic characteristics; they had no [misplaced] yellow or olive hues'. It was inferred that even traces of misplaced yellow found in some individuals in the West Siberian Plain, east of the sympatric region [i.e. 'fulvescens' sensu stricto as used in this web text], are a result of ‘fractional’ introgression of abietinus alleles. Marova et al. (2017) concluded : 'We assume that all specimens with mismatching phenotype and mtDNA – including those caught in the Netherlands – are of hybrid origin'. [Here again, the proviso of correct assignment of morphotype in the first Netherlands study applies.] More disconcertingly, Shipilina et al. (2017) reported that, based on whole genome sequence data, individuals with 'pure morphotoype' and with the corresponding mtDNA could still contain a significant proportion of 'foreign' alleles (i.e. an individual with tristis morphotype and tristis mtDNA could still possess a proportion of abietinus alleles in the genome while an individual with abietinus morphotype and abietinus mtDNA could still possess a proportion of tristis alleles in the genome, the latter being more numerous than the former).
The second Netherlands exercise did not pre-select individuals for analysis on the basis of appearance and also confirmed the occurrence of abietinus as a scarce migrant through the Netherlands, thus tallying with a study by Collinson et al. (2018), which had confirmed the occurrence of abietinus in the UK. The Netherlands data suggested that tristis types occurred at a frequency of between 1% and 2% and abietinus at a frequency of c. 5% of all autumn chiffchaffs. Thus, both taxa were scarce but abietinus now appeared to be slightly more numerous than tristis.
Although the UK and Netherlands genetic studies both confirmed the occurrence of abietinus, albeit as a scarce migrant, both also found that - among the Chiffchaffs trapped during their studies - chiffchaffs carrying abietinus mtDNA were impossible to distinguish from nominate collybita in the field, including via biometrics such as wing-length. Thus, beyond genetic analyses, the diagnosis of abietinus posed a more perplexing task than the diagnosis of tristis. Indeed, as noted above, both Collinson et al. (2018) and van der Spek & de Knijff (2021) suggested that, were it not for genetic differences, the validity of abietinus as a subspecies would be questionable. Such judgements, however, must take into account the features of each taxon as evaluated across its entire range. Judgements must be made at the population level and not on the basis of a subset of individuals of unknown origin. The origins of the abietinus detected in the Netherlands and UK studies are unknown and it is perhaps unlikely that they included individuals from across the full range of abietinus, from Norway to the Urals and south to the Black Sea. Examination of museum specimens has long confirmed that most abietinus are very similar to nominate collybita. Both Ticehurst (A Systematic Review of the Genus Phylloscopus, 1938) and Svensson (Identification Guide to European Passerines, 1992), emphasised their similarity. By implication, many individuals will be impossible to separate from collybita in the field. However, both Ticehurst and Svensson also noted variability in abietinus, with Ticehurst describing the extremes of collybita and abietinus as 'very distinct' (though Ticehurst may have been including within abietinus individuals now designated as menzbieri, a taxon not described until 1937). Svensson noted that the upperparts of abietinus average a little less olive and slightly greyer, while the underparts are a little less yellow. More critically, individuals in the east of the range he described as displaying more grey and buff and individuals in the NE as sometimes paler overall and with whiter underparts.
Dean & Svensson (2005) also included 'variants among all the subspecies' in their list of sources of greyer individuals and 'greyer' variants have been documented even among nominate collybita. However, significantly greyer Chiffchaffs confirmed as bearing nominate collybita mtDNA remain very few. Collinson et al. (2018 ) published a photo (plate 276 therein) showing a Chiffchaff with grey and white aspects to its plumage yet nominate collybita mtDNA. However, based upon the photo, its appearance overall seems aberrant, with the upperparts exhibiting an almost 'slatey' tone. It does not look like tristis nor a a 'grey-and-white Bonelli's-like' Chiffchaff while neither does it match the appearance of any of the southern forms.
Traditionally, it has been considered that the southern forms of Chiffchaff, brevirostris, caucasicus and menzbieri, would be unlikely to reach western Europe. Recent studies, however, have indicated that this assumption is false. Further comments on the southern subspecies are provided below.
It is now clear that diagnosis of Chiffchaffs must always be based upon careful evaluation of plumage and vocalisations and that all taxa must be taken into consideration.
Various comments have been made relating to the likelihood of hybrids and intergrades between tristis and abietinus reaching Britain. These comments infer that, given the vast range of tristis in comparison with the relatively narrow hybrid zone, the likelihood of individuals reaching Britain from the hybrid zone is small. This implies that individuals reach western Europe with equal likelihood from across the entire range, from the Urals to the Kolyma River. This is implausible and it seems more likely that regions closer to western Europe, including the hybrid zone, will be the source of the majority of tristis-candidates encountered in western Europe (see Dean 2013).
In December 2024, a paper was published by Dufour et al. in Ibis, investigating the use of age ratios to examine movements of Yellow-browed Warblers P. inornatus and Siberian Chiffchaffs (treated by them as a separate taxon from Common Chiffchaff, thus P. tristis). The study involved trapping 324 inornatus and 117 tristis in Sweden and France, the tristis being identified using call and the colouration criteria as listed above in 'The defining features of 'classic' Siberian Chiffchaffs. Among much else of interest, 56 tristis candidates trapped in France were genotyped using feathers collected during the captures. As well as mtDNA, SNPs were analysed in order to detect evidence of hybridization and introgression. Hybrid scores were allocated on a range of 0 to 1, with any score below 0.05 regarded as pure abietinus and any score over 0.95 regarded as pure tristis. The paper includes the following regarding tristis candidates reaching Europe:
'Interestingly, genomic results showed that all our sampled birds were composed of dominant tristis ancestry but that a significant proportion were intergrade individuals (i.e. with a hybrid score < 0.95), suggesting past-hybridization and introgression. This would suggest that at least some of the birds breed in the west of the tristis distribution close to the contact zone between abietinus and tristis (Shipilina et al. 2017). Further more, this raises new hypotheses concerning the emergence of new migration routes, with hybridization possibly leading to the introgression of genes or genomic regions involved in the orientation of migration (but see Delmore & Irwin 2014).'
Individuals diagnosed as intergrades with a hybrid index of <0.95 numbered 19 of the 56 analysed genetically, i.e. 34%. From Fig. 3 in Dufour et al., it can be deduced that about 11% involved a hybrid index of between 0.88 and 0.65.
In Britain, White & Kehoe (2021) estimated that 2968 tristis were recorded between 2008 and 2019. Were the results from individuals trapped in France translated to Britain, then a proportion of 34% would imply that over 1000 involved some degree of introgression from abietinus. However, if matching the French results, in many of these the level of introgression would be relatively low. A proportion of 11% would translate to about 320 of the 2968 tristis candidates estimated by White & Kehoe for 2008 to 2019. Courtesy of Paul Dufour, I have been able to examine all the photographs and their 'hybrid indices'. Some suggest slight 'fulvescens' traits but these images include some where the overall colour-balance of the images indicates that colour artefacts are involved. More confidently, among the high quality portrayals, of 33 individuals evidently matching fully the criteria for ‘classic’ tristis, 23 were genotyped as 'pure' tristis (70%). However, 10 (30%) were genotyped as ‘backcrosses’.
This supports the conclusion of Shipilina et al. (2017), that some individuals with perfect tristis morphotype can nevertheless be intergrades. Further, the results of Dufour et al. indicate that such intergrades are reaching Europe. Dufour et al. suggest that the use of geolocators could help to reveal the origins and movements of tristis reaching Europe.
[Note on the level of introgression. Among the tristis morphotypes analysed by Dufour et al., the 'lowest' hybrid index (i.e. the lowest level of tristis in the genome) was 0.64 and this individual was interpreted as an F2 (second generation) hybrid. Thus, although backcrosses were numerous, comprising 34% of tristis morphotypes analysed, no first-generation (F1) hybrids were encountered. The genetic composition of individuals within the hybrid zone will include F1 hybrids, which are confined to the 'current' year, but also a presumably increasing proportion of backcrosses, as these will aggregate over successive generations. Commentary from a geneticist on the likely spectrum of introgression levels within the hybrid zone is necessary. However, the results of Dufour et al. would seem to imply that most individuals with mixed alleles originated from the region immediately east of the hybrid zone or that, within the hybrid zone, backcrosses now significantly outnumber the number of F1 hybrids produced in a given year.]
Although many questions remain relating to tristis-candidates reaching Britain and western Europe, Shipilina et al. (2017) have provided genetically-based evidence for the origins of morphological variation observed among tristis in the region of the Urals while also confirming that, in contrast, the appearance of individulas in the Yenisey region and Yakutia is consistent. They concluded that the taxa tristis and abietinus hybridize extensively in the sympatric region and genetic admixture is the principal origin of Chiffchaffs which deviate in appearance from 'classic' individuals. Dufour et al. (2024) have demonstrated that tristis candidates trapped in France 'were composed of dominant tristis ancestry but that a significant proportion [34%] were intergrade individuals'. An implication was that 'at least some of the birds breed in the west of the tristis distribution close to the contact zone between abietinus and tristis'. Both these studies also indicated that individuals with perfect tristis morphotype may yet be intergrades. What are the implications of these genetic results for field identification of Siberian Chiffchaff?
It seems that both appearance and vocalisations are controlled by a limited area of the genome (Shipilina et al. 2017, Talla et al. 2017), so that even an individual with the appearance and song of tristis may yet harbour ‘foreign alleles’. Shipilina et al. note that: ‘analysis of genomic composition of birds within the sympatric zone showed that several individuals express diagnostic plumage characters and perform diagnostic song despite harbouring the foreign mtDNA type and/or a considerable proportion of foreign nuclear alleles’. Full trait data were available for only a limited sample but indicated that abietinus morphotypes carrying tristis mtDNA were significantly more numerous than tristis morphotypes carrying abietinus mtDNA (among mixed populations studied by Marova et al. (2017), 56.7% of abietinus phenotypes cf. 4% of tristis phenotypes harboured a contrary mtDNA haplotype). Despite this imbalance, Marova et al. (2009, 2013) reported an individual with tristis morphotype and tristis song yet carrying abietinus mtDNA. These statistics on mtDNA mismatches have been interpreted as implying that a Chiffchaff which ‘looks like’ tristis and 'sounds like' tristis has a reasonably high probability of actually being a thoroughbred tristis. However, these statistics relate to mtDNA and overlooks the finding by Shipilina et al. that even individuals with matching morphotype (appearance) and mtDNA may yet harbour 'a significant number of foreign alleles' i.e. the nuclear genes are mixed. What the findings of Shipilina et al. indicate is that a Chiffchaff which ‘looks like’ tristis has a reasonably high probability of having tristis mtDNA - but it may still harbour a proportion of abietinus genes elsewhere in the genome. Subsequent genetic studies by Dufour et al. (2024) reinforced the conclusion that tristis morphotypes may yet be intergrades.
Also, in practice, in the field, matching morphotype and mtDNA begs the question of what is meant by ‘looks like tristis’? In the context of the paper by Shipilina et al., ‘looks like tristis’ means a total absence of misplaced yellow (as per the original ‘Svensson criteria’). Unfortunately, a complete absence of such yellow can only be guaranteed by examination in the hand. For this reason, the Swedish RC, for example, at one time would accept only trapped individuals. In the field it is impossible to confirm an absolute absence of misplaced yellow (or olive).
Similarly, what is meant by 'sounds like tristis'? While careful examination of sonograms will reveal 'mixed' singers, in the field 'mixed' song can be very difficult to distinguish from standard tristis song (Marova et al. 2013; & pers obs), with the more varied tristis-style notes determining the overall character of the song and readily masking a minority of abietinus notes.
Compounding the practical difficulties, in the studies by Marova et al. and Shipilina et al., pure tristis song was relatively frequent among individuals with 'intermediate' appearance (i.e. with some yellow streaking), which may easily be assessed as 'looks like tristis' in the field, while individuals with 'pure' tristis morphotype (determined in the hand) were frequently 'mixed singers'. These studies did not involve call but van den Berg et al. (2009) reported that tristis-style Chiffchaffs encountered on migration in Kazakhstan exhibited 'wide plumage variation' yet called and sang exclusively like tristis (though sonograms were not provided). At the time, this was interpreted as indicating extensive plumage variation among thoroughbred tristis but the findings of Shipilina et al. proffer a rather different interpretation : that 'wide plumage variation' was a sign of genetic admixture and that hybrids may call (as well as sing) like tristis.
Shipilina et al. (2017) suggested that their results brought into question the feasibility of identifying abietinus and tristis on the basis of appearance. So, do we return to an approach which disbars field identification completely or even demand 'whole genome' sequencing of candidate tristis? I am no geneticist but I would argue not. While the level of genetic mixing in the sympatric zone around the Urals (colloquially dubbed here as the ‘riphaeus’ zone) is relatively high, the level of introgression among many individuals with very limited traces of yellow found in the allopatric West Siberian Plain (here the ‘fulvescens’ zone sensu stricto) is likely to be low (‘fractional’ to use the word employed subsequently in the Shipilina et al. paper). In this region, the two forms are basically allopatric. It is suggested here that individuals with such 'fractional' introgression from the allopatric West Siberian Plain surely remain designated reasonably as Siberian Chiffchaffs? Lars Svensson, in Dean & Svensson (2005), has suggested that 'fulvescens' from the West Siberian Plain (sensu stricto) may be best accommodated within tristis. See also Appendix 3 for further - but tentative - comments on the genetic issues.
The criteria adopted by the BBRC’s ‘tristis panel’ in 2008 were described in detail by Stoddart (British Birds, 2008, 101: 165-166) and Dean et al. (British Birds, 2010, 103: 320-338) and reflect this interpretation. The identification criteria they recommended were formulated in 2008 (nine years prior to the 'whole genome' findings of Shipilina et al. 2017) but the panel concluded that individuals which matched the ‘Svensson criteria’ under close and careful scrutiny in the field were acceptable, thus accommodating the likelihood that, if examined in the hand (or in high resolution digital photos), some would reveal marginal 'fulvescens' traits. However, individuals with field-evident levels of yellow were excluded by the panel’s criteria, as their origins were uncertain and could well include F1 hybrids and early backcrosses from the sympatric ('riphaeus') region around the Urals. It was recognised that some backcrosses from the sympatric zone would overlap in appearance with those from the allopatric zone exhibiting just trace levels of yellow but a ‘pragmatic’ approach to field identification was advocated.
The panel’s criteria were pragmatic but exacting and explicitly excluded individuals with field-evident levels of misplaced yellow and olive, which places them on the wrong side of the 'riphaeus' line. Also excluded were the still-contentious ‘grey and white Bonelli’s-like’ Chiffchaffs, which not only have traces of yellow but also lack the characteristic ‘tan-brown and buff’ hues of classic tristis.
The evident proviso for the 'pragmatic' approach advocated by the panel is that a majority of tristis morphotypes reaching the UK and western Europe are thoroughbreds or have only very low levels of introgression. To some extent this will reflect their geographical origins. The recent genetic study by Dufour et al. (2024) has shown that, among among tristis candidates trapped in France, 34% showed at least some level of introgression from abietinus (see paragraphs above on genetic studies) but for most individuals, the level of introgression was relatively low. However, several perfect tristis morphotypes proved to be intergrades. Further research into the origins of tristis-candidates reaching western Europe are planned (Talla et al. 2017, Dufour et al. 2024). In the interim, the phrase 'showing the characters of Siberian Chiffchaff ' may be appropriate for records of individuals identified solely on the basis of field observations.
'Relaxed criteria', embracing putative 'Siberian Chiffchaffs' with field-evident levels of misplaced yellow and olive, have been advocated by some in recent years, based upon claims that hybridisation was rare and that the number of tristis-candidates reaching Europe from the overlap zone would be insignificant. Such claims are now shown by genetic studies to be misguided. Hence, birds with field-evident levels of misplaced olive and yellow or with conflicting vocalisations, must remain excluded. It must also be acknowledged that accepting individuals on the basis of full compliance with the tristis morphotype will still involve a margin of error. Needless-to-say, anomalous Chiffchaffs 'still have a story to tell' and should be documented and not ignored. It should not be forgotten, also, that the statistics available so far suggest that, although difficult to diagnose with certainty, many thoroughbred tristis reach Europe. 'Classic' appearance and 'classic' vocalisations remain the credentials by which candidates should be assessed, albeit not failsafe.
Dean & Svensson (2005) suggested several possible sources of 'greyer' Chiffchaffs. They noted that, as well as greyer examples of both abietinus and tristis, options to be considered included hybridization between these two forms ('riphaeus' types), individual variation among all the races and the possibility of Chiffchaffs in some part of the range consistently combining greyer livery with a tristis-like call. As noted above in respect of the last of these options, recent studies confirm that such Chiffchaffs do exist, in the SE of the species' range : notably menzbieri and caucasicus (current data suggest that brevirostris is a browner form).
Southern forms confirmed well outside traditional ranges.
The genetic studies of Raković et al. (2019) implied a much-enlarged breeding range for menzbieri, extending from the traditionally recognised range in Turkmenistan and NE Iran, continuously through the Elborz mountains of Iran and into southern Armenia. In the UAE, Chiffchaffs with menzbieri DNA were trapped in March 2019 (one) and December 2020 (two) : Motteau et al.( 2022). As these comprised three of four Chiffchaffs genetically analyzed (the other being abietinus), the implication drawn was that menzbieri is a regular component of the Chiffchaffs wintering in the UAE and probably elsewhere in the Middle East. An extensive ringing and genetic study by Babbington et al. (2024) detected a small number of brevirostris /caucasicus in Saudi Arabia during February and March, the first confirmed records of this subspecies-pair. This publication includes informative photographs of Chiffchaffs with mtDNA signatures of brevirostris/caucasicus, abietinus and collybita. No menzbieri were found but, as birds were not trapped during mid-winter, further studies were required to determine the range of subspecies occurring at that season.
See <here> for discussion of the appearance and calls of Chiffchaffs wintering in Kuwait and <here> for discussion of the appearance and vocalisations of Chiffchaffs breeding in the Elborz Mountains of Iran and in Armenia.
Southern subspecies of Chiffchaff proven to reach western Europe
For a long time it was considered that SE forms were unlikely to reach NW Europe but, during 2015, Chiffchaffs with brevirostris/caucasicus mtDNA (the two having long been deemed indistinguishable by mtDNA) were trapped in the Netherlands (two) and Germany (one) : van der Spek & de Knijff (2021).
In a paper published in Ibis in 2025, van der Spek et al. reported that a new protocol developed by Peter de Knijff had enabled the mtDNA of brevirostris and caucasicus to be distinguished. This new technique was employed to analyse the mtDNA of six brevirostris/caucasicus individuals by then recorded in western Europe. The results indicated that an individual in Germany in May2022 carried brevirostris mtDNA while five caucasicus were recorded during late autumn and early winter from the Netherlands and Germany. The authors concluded that caucasicus might be overlooked in Western Europe, rather than an exceptional vagrant.
[Note: Photographs were published by van der Spek et al. of the individuals assigned to caucasicus. Figures 3 & 4 there show individuals with reduced olive and yellow in their upperparts and relatively white underparts, thus similar to Chiffchaffs observed in Armenia and Iran (see <here> ) and also resembling the greyer individuals assigned to abietinus illustrated above. Perplexingly, however, two of their photographs are morphologically at odds with the appearance of caucasicus and look closer to tristis. Indeed, one (in their Fig 2) is morphologically a 'classic' tristis.]
The conclusion from these Middle East and European studies is that brevirostris and caucasicus can reach Europe and raise the prospect that menzbieri could do so also. Thus, a comprehensive approach to the diagnosis of tristis must give regard to the appearance and calls of the more-southerly races.
Call will certainly provide invaluable guidance when diagnosing Chiffchaff taxa, at least when an individual calls frequently and with a consistent call. However, many candidate tristis call infrequently (even not at all) while call will not exclude the possibility of hybrid origin. Also, 'alternative' and 'plastic' calls of collybita / abietinus may simulate or suggest the standard call of tristis and vice versa and it is now clear that such calls are not confined to juveniles in the immediate post-breeding period, as has been claimed in the past. Some individuals combine 'variant' calls with plumage anomalies and thereby raise further questions about their genetic origins. Adding to these complexities, there have been several instances of Chiffchaffs with plumage features firmly associated with tristis yet calling persistently like collybita/abietinus. During the winter 2013/2014, several such individuals were photographed and/or video'd and also sound-recorded, placing their combination of plumage and call beyond question (see 'Case Study 3'). Finally, it is now being shown that the southern subspecies of Chiffchaff can reach Europe (see above) and these taxa have calls which can sound very similar indeed to tristis. Sonograms show that there are distinctive shapes in some instances but that in other cases even sonograms display a tristis-like profile (see <below> and 'Chiffchaffs in Iran and Armenia'). Earlier assertions that calls are a failsafe guide to distinguishing Chiffchaff taxa, and render detailed plumage evaluation unnecessary, are now shown to be misguided. Diagnosis must always be founded on a careful evaluation of both plumage and vocalisations. The topic of vocalisations is examined in more detail below.
The characteristic call of tristis ► (here recorded in Russia) is a near-monosyllabic ‘eeep’ (or ‘iiihp’ in Scandinavian renditions), predominantly even in pitch but descending and fading slightly at the end, which produces a plaintive and slightly 'off-key' quality.

Figure 1.
In sonogram, note predominantly even pitch ('flat' and horizontal trace) with just a slight downturn at the end.
The cd 'Calls of Eastern Vagrants' (Jännes 2002) also includes a particularly good and monotone example, recorded in the wintering grounds in India. The sound recalls a less sibilant and less 'tinny' version of a Dunnock's call or a higher-pitched less piping variation of a Bullfinch's call. This call is very different from the normal, distinctly rising ‘hüit’ of collybita and abietinus, and is an important component in any identification of a potential tristis.
The song of tristis ► (here recorded in Russia by Antero Lindholm) is a fluent warble of closely-spaced and near-disyllabic notes. Sonograms of the song show characteristic notes with ascending modulations, which are absent from the typical song of collybita and abietinus. These ascending and disyllabic-sounding notes generate a characteristic rising and falling character, much more varied and liquid than western chiffchaff song. A typical sequence might be transcribed as: ‘chivvi-tee, chooee, chivvi-tee, chooee-tee, chivvy’. While tristis song does contain occasional notes with a descending pitch overall (and which can recall abietinus/collybita notes in structure), such notes are in a minority and are absent entirely from some tristis songs (and perhaps from the repertoire of some individuals). Also, they often start with a short rising element and have a distinctly narrower bandwidth than true abietinus/collybita notes. See discussion in Lindholm (2008) for details of note structure in the songs of abietinus and tristis and also a comprehensive examination of 'mixed singers' (see also 'Mixed Singers' below).
Ivanistkii et al. (2020) have reported that the short, dry 'tret' calls, which Chiffchaffs interpose between the strophes in their songs, also differ between the various taxa of the chiffchaff complex (including here the 'Mountain Chiffchaffs' lorenzii and sindianus). The exception was that the 'tret' calls of collybita and abietinus were indistinguishable and there was a proviso over brevirostris, which (at least during their study) used such calls relatively infrequently. Intriguingly, their findings included that the 'tret' calls of tristis differed between central Siberia and NE Yakutia.
Although adults of all races have 'standard' contact calls, which they employ consistently under most circumstances, there are also so-called 'plastic' and 'alternative' calls. Such variant calls are employed regularly by juveniles in autumn while, in the cases of collybita and abietinus at least, alternative calls are also employed by adults under certain circumstances and in certain years (for reasons which are not yet understood). So-called 'plastic' calls are associated with juveniles which are deemed to be learning their calls. At this early stage, collybita may give a call which is very reminiscent of tristis, being both even in pitch and monosyllabic. Here ► is a composite recording which includes, first, a juvenile collybita (Warks, July 2013) and then a tristis (Warks, January 2013). In such direct comparison, the call of the juvenile collybita is marginally less even and rather more 'fuzzy', less 'pure', than the tristis call but they are in general rather similar. Below is a sonogram comparing the pitch and structure of the two calls (Fig. 2.).
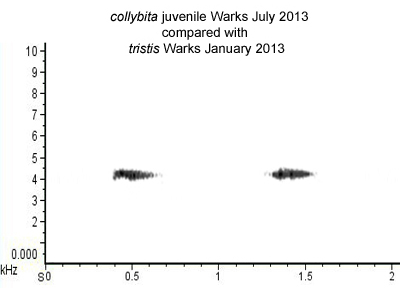
Figure 2.
See also 'The Sound Approach to Birding' (Constantine & The Sound Approach, 2006), page 91, and listen to the call at 0:16 - 0:18 in their recording of Common Chiffchaff on CD1 track 86.
It is sometimes claimed that such 'plastic' calls are unlikely to be heard after early September, well before the arrival time in western Europe of tristis. However, it is now known that 'plastic' calls can persist into November, at least. Although rarely as similar to tristis as the 'plastic' call illustrated above, confusion can also arise between the classic call of tristis and 'alternative' calls given at times by abietinus and collybita at any time of the year. Such 'alternative' calls are very different from the standard, rising ‘hüit’ or 'hweet' call and can easily lead to confusion. Among documented 'alternative' calls, perhaps the most frequent reference is to a call usually transcribed as 'sweeoo', which rises then falls rather sharply in pitch, creating a disyllabic sound, while the sonogram has the shape of a circumflex or an inverted 'V'. It can recall one call of Thrush Nightingale Luscinia luscinia. This type of call has been associated with 'eastern abietinus' (see for example Jännes 2002) but such a call is also to be heard from collybita and western abietinus, at any time of the year but especially during the late summer and autumn. They have been heard even during the winter in the UK (pers obs) and also in Spain, from birds with the appearance of western forms (Copete & Armada 2004). Additionally, in some years they have been heard commonly in spring from adults, in the UK and elsewhere in Europe (see separate article on this topic).
Despite the disyllabic quality, the fact that the 'sweeoo' type of call is rather abrupt, and has start and end frequencies which are closer than in the evidently rising 'hweet' call, means that even well-inflected 'sweeoo' calls can have a 'flatter' sound than the familiar 'hweet' call.
Here is a composite recording ► which compares the 'sweeoo' call of a collybita (UK) with the classic 'eeep' call of tristis (Russia). A sonogram of the two calls is below (Fig. 3):
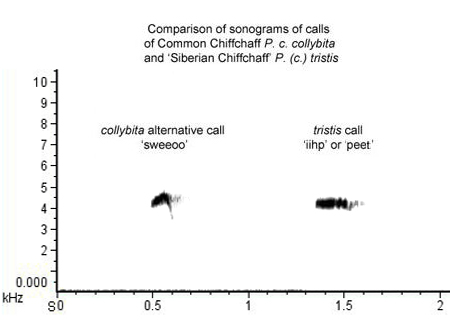
Figure 3.
In such a direct comparison, the 'alternative' call of collybita is quite abruptly and strongly inflected and has a 'brighter' timbre, while the call of tristis is straighter and is more plaintive and slightly 'off-key'. However, it is easy to understand how the two might be confused when heard in isolation, at distance, or by someone not thoroughly familiar with the call of tristis.
Conversely, not all calls given by tristis and/or 'fulvescens' match the ideal or classic 'straight' structure described above. Calls attributed to tristis may also have a clear rise and fall in pitch on occasions, and could be transcribed as a shallow 'sweeoo' rather than a truly evenly-pitched and monosyllabic 'eeep' or 'iiihp'. Here is a composite recording ► which compares two calls attributed to tristis: a classic, straight 'eeep' call from Russia and a shallow 'sweeoo' call from India. A sonogram of the two calls is below (Fig. 4).

Figure 4.
Generally, compared with the 'sweeoo' calls of collybita and abietinus, the shallow 'sweeoo' call of tristis / 'fulvescens' has a much more gentle transition from rising to falling pitch and a distinctly 'shallower' frequency range and lower peak frequency. The sonogram has the shape of a shallow inverted 'u' rather than an inverted 'V'. See Antero Lindholm & Anneka Forsten's website for a variety of such calls.
Here is a composite recording ► which compares a 'sweeoo' call given by a nominate collybita in Warwickshire in July 2011 (my own recording) with a 'sweeoo' call given by a tristis / 'fulvescens' in India in December 2007, recorded by Antero Lindholm. A sonogram of the two calls is below (Fig. 5).
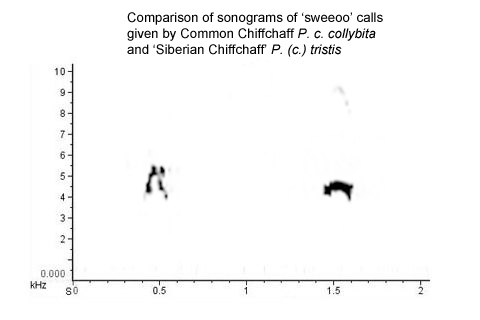
Figure 5.
This direct comparison and the sonograms illustrate the relatively abrupt transition of the collybita call and the smoother transition of the tristis / 'fulvescens' call, together with the differences in peak frequency and frequency range, which create a different timbre. Yet, both are clearly ‘sweeoo’ calls to the ear and could well be confused if heard in isolation or in the imperfect acoustic conditions which will generally attend field encounters. For example, listen to the recording and examine the sonogram of an abietinus/collybita style 'sweeoo' call given by a 'mixed singer' Chiffchaff in Warks in March 2014). The vocabulary of other forms of Chiffchaff is varied and tristis is unlikely to provide an exception. Listen here ► and here ► to series of recordings from October 2009 by Maarten Wielstra, involving a Chiffchaff in the Netherlands which looked and sang like tristis. Some of the calls in the series are decidedly different from the classic, evenly-pitched 'iiihp' or 'eeep' call of tristis.
Alternative calls themselves vary, with some 'sweeoo' type calls given by collybita and abietinus being truncated and less-emphatically inflected, thus resembling the 'plastic' calls of younger juveniles and converging with the shallow 'sweeoo' call given by some tristis (both readily sounding more like a monosyllabic 'swee'). Some individuals give such calls persistently. The indications are that 'sweeoo' type calls can be given by all forms of Chiffchaff and provide the most evident case of call convergence.
As well as the regularly encountered 'standard' and 'alternative' calls, Chiffchaffs of all taxa occasionally utter 'wayward' or 'eccentric' calls, which differ from the established calls which are employed more systematically. As these too may sometimes suggest the standard calls of another Chiffchaff taxon, they must also be considered when encountering an anomalous call. Generally, such 'eccentric' calls will not be used persistently and will also lack consistency in their structure and 'sound', while 'standard' calls will also be interspersed in 'eccentric' call-sequences.
The calls of Chiffchaff races in the Middle East and the Caucasus
The calls of Chiffchaffs encountered in Turkey (brevirostris), the Caucasus (caucasicus) and in northern Iran and Kopet Dagh in Turkmenistan (menzbieri) are relatively flat and may resemble or even match the typical call of tristis (e.g. Copete & López 2013; Lars Svensson in litt., Dubois & Duquet 2008., pers obs). Indeed, texts asserting that the call of tristis is diagnostic, irrespective of the caller's appearance, have led to some visitors to the Middle East from western Europe misidentifying southern forms of Chiffchaff as tristis.
The taxonomy of these forms is still debated. Currently, they are accorded subspecific rank but, historically, they have also been subsumed within abietinus (and menzbieri sometimes subsumed with tristis / 'fulvescens' - see Appendix 2). More recently, based upon their evaluations of morphology, Shirihai & Svenssson (2018) have advocated that brevirostris be subsumed into collybita and caucasicus into abietinus (with reservations expressed too over recognition of menzbieri). In relation to calls, if caucasicus (and perhaps menzbieri too) were to be subsumed into abietinus, then complications would be resurrected (1) over the vocabulary of 'abietinus' across the resulting much-expanded range, with both the widely-recognised rising 'hweet' of northerly populations and the tristis-like calls of southerly populations being deemed to be within the vocabulary of 'abietinus'. Even more confusingly, if brevirostris were to be subsumed into collybita, then the vocabulary of collybita too would be deemed to include a tristis-like call.
However, while brevirostris may be less distinct in appearance from collybita and abietinus, the forms found further east in this region are evidently greyer above and whiter below compared with typical abietinus and have relatively little yellow on the underparts i.e. in the regions embracing the ranges of caucasicus and menzbieri as now perceived. See 'Other sources of 'greyer' Chiffchaffs' above. All three taxa differ in mtDNA from abietinus. In conjunction with their relatively 'flat' and tristis-like calls, taxonomic recognition of all three southern forms seems warranted.
More research into the calls (and range of variation and overlap of calls) of the southern subspecies is required. However, recent studies indicate that, while the calls of brevirostris, caucasicus and menzbieri can all strongly recall tristis to the ear, there are subtle differences to be found in sonograms. At least brevirostris ad menzbieri may employ calls which begin on an even pitch, very like trsstis, but then lead into a 'terminal arch' (i.e. a flat introduction, then a subtle 'sweeoo' structure). There is evidence of a degree of call variation, however, with a slightly down-sloping call also associated with brevirostris while very flat/straight (and extremely tristis-like) calls have been recorded form individuals deemed to be caucasicus and menzbieri based upon range. Whether these variations are individual variation, seasonally influenced or even geographically-based remains to be elucidated (given the history of Chiffchaff subspecies, it is even conceivable that a cryptic subspecies has yet to be diagnosed within the complex).
See <here> for discussion of these issues and the appearance and vocalisations of Chiffchaffs breeding in the Elborz Mountains of Iran and in Armenia. That text includes sonograms of Chiffchaffs attributed to menzbieri and comparative sonograms from Helbig et al. (1996) and Clement et al. (1998), which include calls attributed to caucasicus, brevirostris and menzbieri. Similarities and potential subtle differences are explored. For discussion of the calls of enigmatic Chiffchaffs wintering in Kuwait see <here>.
See <above> for comments on southern forms encountered outside their traditional ranges.
[Footnote (1) : Across its European range the standard call of abietinus is identical to that of collybita, contrary to some earlier assertions that the call of 'eastern abietinus' - meaning in a European context - involved or included a tristis-like call. This error probably arose when European birdwatchers first began to visit the Middle East and heard tristis-like calls which they assumed were being given by migrant abietinus. This is further confirmation that call alone is not a sufficient basis for assigning Chiffchaffs to subspecies. Within the range of abietinus as defined by its current taxonomy, the standard call of abietinus is consistent. ]
Conflicts between morphology and call type
The paragraphs above relate primarily to calls used by Common Chiffchaff which can cause confusion with the typical call of tristis. An even more perplexing issue involves Chiffchaffs with plumage features associated with tristis/'fulvescens' yet persistently employing fully-crystallized collybita/abietinus calls. These have not been isolated, hesitant calls with the character of 'plastic' calls but prolonged series of fully-developed collybita/abietinus calls. Several of these individuals occurred in Britain during the winter of 2013/2014 and were observed over several weeks. They used such collybita/abietinus calls persistently and not as a fleeting departure from the typical tristis call. Are such anomalous callers the result of cultural call 'copying' among perceived competitors (and if so what does it imply about their geographical origins) or do they have a genetic origin? Do call anomalies have a cultural origin or are they too a sign of genetic introgression? This intriguing issue is explored further, with examples, in 'Case Study 3 : conflicting morphology and vocalisations'.
The origins of 'mixed singers'
As well as individuals with a conflict between morphology and call, Chiffchaffs using songs incorporating notes of both Common Chiffchaff and Siberian Chiffchaff are sometimes encountered in the UK and western Europe. These can be genuine 'mixed singers' (with an intimate mixture of notes from the two taxa in a single song strophe) or 'song switchers' (giving full strophes of the two taxa alternately). Song anomalies have been associated with genetic admixture (see Shipilina et al. (2017) but, as song is deemed to be primarily learned in oscine passerines, some claim that such mixed song could be the result of 'copying' by thoroughbreds. No doubt both mechanisms could be involved in individual cases. However, in each case the implication is that the individuals involved originate from the 'overlap' zone in and around the Urals. This is clearly the case for 'hybrids' but also for 'mixed singers', as individuals raised farther east in Siberia will not be hearing abietinus song during the critical song-learning phase. Again, see 'Case Study 3'.
None of this should undermine the significance of the classic tristis call, which still provides an important element in identification. However, to regard call as an unfailing key is over-simplistic and diagnosis of tristis should always embrace both vocal and plumage features.
In summary:
The
classic evenly-pitched, monosyllabic and somewhat mournful call of
tristis ('eeep' or 'iiihp', with a flat shape in a sonogram, thus:
![]() ) is a strong indicator of taxon but it is essential
that the call is heard clearly and that the observer is thoroughly familiar
with the call of tristis. Ideally, the call should be confirmed by
sound recording and sonogram.
) is a strong indicator of taxon but it is essential
that the call is heard clearly and that the observer is thoroughly familiar
with the call of tristis. Ideally, the call should be confirmed by
sound recording and sonogram.
The normal call of collybita and abietinus is a 'bright' and rising 'hweet', quite different from the standard call of tristis.
However, so-called 'alternative' calls are also given regularly and consistently by collybita and abietinus and have a rising and falling frequency and a rather disyllabic sound, usually transcribed as 'sweeoo'. Despite their disyllabic quality, such calls are rather abrupt and the fact that their start and end frequencies are close means that they can have a 'flatter' sound than the familiar, rising 'hweet'. Correspondingly, such calls can be confused with the classic, straight tristis call.
Emerging evidence suggests that the call of tristis does not
invariably match the idealised, evenly-pitched and monosyllabic 'eeep' or 'iiihp'
but may rise and fall in frequency slightly. Thus, tristis too may
use a 'sweeoo' variant call, though this is shallower than the 'sweeoo' call
of abietinus/collybita. The transition from rising to falling pitch is usually more gradual (less
abrupt) than in collybita/abietinus, the peak frequency is
lower and the frequency range narrower (more compressed). Thus, in a
sonogram, the shapes of the calls recall a shallow inverted 'u'
for tristis and an inverted 'v' for collybita/abietinus
(i.e.
![]() ).
).
Thus, there is some convergence in structure and 'sound' between such variant 'sweeoo' calls, with a 'two-way' potential for confusion. A recording and sonogram will provide structural clues but, clearly, attributing such calls to a given taxon by ear alone can be problematic.
Chiffchaffs of all taxa will occasionally employ 'eccentric' calls which may suggest the standard call of another Chiffchaff taxon. Generally, such calls are used intermittently, or the call-structure lacks consistency, so that prolonged listening and recording will reveal its 'eccentricity'.
More perplexing are occasional individuals with plumage traits which are associated with tristis (e.g. a deficiency of yellow and olive in the plumage) yet employ persistent series of perfectly formed abietinus/collybita calls. Some have proved additionally to be 'mixed singers' (see Case Study 3'). A cultural origin has been suggested for such vocal anomalies (leading to one taxon 'copying' from another) but studies of tristis and abietinus in their sympatric zone have revealed an origin in genetic admixture (see Shipilina et al. (2017).
It should be borne in mind also that the calls of the more southerly forms brevirostris, caucasicus and menzbieri can be similar or even indistinguishable by ear from that of tristis (e.g. Lars Svensson & José Luis Copete in litt.; Dubois & Duquet (2008); pers obs). Chiffchaffs with the mtDNA of caucasicus/brevirostris have been trapped in the Netherlands (two) and Germany. Thus, a tristis-like call does not guarantee the identity of an enigmatic Chiffchaff. (See <here> for discussion and sonograms of Chiffchaffs in Iran and Armenia, where the significance of calls with a 'terminal arch' is explored.)
Variant, anomalous and convergent calls illustrate the importance of assessing both vocalisations and morphology of any candidate 'Siberian Chiffchaff'.
In 2007 the BBRC co-opted a 'tristis panel' to examine all reports of 'Siberian Chiffchaff' in the UK during the 'sample' year of 2008. The panel examined all individuals using the traditional 'Svensson criteria' for classic tristis, while taking into account the form's characteristic 'tan-brown and buff' hues. Also, allowance was made for individuals which were clearly very like 'classic tristis' but exhibited very limited 'fulvescens' characters (generally imperceptible in the field and often detected only in good, close-up photographs). To insist on an absolute absence of 'additional' yellow and olive would, in effect, exclude field identification of tristis completely, as the absolute absence of these 'misplaced' hues can only be confirmed in the hand. To exclude field identifications would, in turn, undermine the objective of establishing a measure of the true status of Siberian Chiffchaff in Britain. Further, as discussed above and in the Appendices, individuals with very slight 'additional' yellow and olive (i.e. with limited 'fulvescens' traits) are compatible with chiffchaffs from the West Siberian Plain and beyond the true zone of overlap with abietinus. Although slight introgression across the West Siberian Plain is likely, the panel concurred with the suggestion by Ticehurst (1938) and by Lars Svensson (in Dean & Svensson 2005) that individuals with such restricted 'fulvescens' traits are best incorporated with tristis (though with no formal taxonomic implications in the term 'fulvescens'). Conversely, 'riphaeus'-like individuals (with more evidently intermediate plumage), individuals with anomalous calls, and 'Bonelli's-like' grey-and-white individuals (sensu Dean & Svensson 2005), were excluded by the panel's criteria, as the origins of such individuals remained equivocal. While questions remain to be answered, a clearer picture of the status of tristis (here including 'fulvescens' sensu stricto) in the UK emerged. Full details were published in: Dean, A., Bradshaw, C., Martin, J., Stoddart, A. & Walbridge, G. 2010. The status in Britain of 'Siberian Chifchaff'. British Birds 103: 320-338. The panel's investigations demonstrated that the 'Siberian Chiffchaffs' meeting these criteria reach Britain in significant numbers. For 2008, reports of 57 individuals were submitted to the panel, of which 49 (86%) were accredited. Applying this endorsement rate to all known reports in Britain during 2008 suggested that around 120 Siberian Chiffchaffs may well have occurred during the year. Thus, Siberian Chiffchaff was confirmed as a scarce rather than rare visitor, a conclusion confirmed by analyses for subsequent years that have been published in the 'Report on Scarce Migrants' published periodically in British Birds. The tristis panel's report also examined in detail the issues of morphology, vocalisations, genetics and hybridization (as known at that time); the seasonal and geographical distributions of Siberian Chiffchaffs reaching Britain; comparative data from other European countries; and the numbers of Siberian Chiffchaffs reaching Britain in comparison with those of Yellow-browed and Pallas's Leaf Warblers. Reference should be made to the panel's publication by anyone wishing to explore the topic further.

© A. R. Dean
Plate 10. Apparent 'Siberian Chiffchaffs' with enigmatic vocal and plumage attributes
A case-study from Worcestershire, by Andrew Warr, winter 2014/2015 (external link)
For much useful discussion, information, clarification and / or provision of excellent photos, I am indebted to Jem Babbington, Niclas Backström, Staffan Bensch, Colin Bradshaw, Martin Cade, Oscar Campbell, Richard Chandler, Martin Collinson, Greg Conway, José Luis Copete, Gary Crittenden, Philippe Dubois, Daniel Duff, Paul Dufour, Martin Garner, Dave Hutton, Gabriel Jamie, Hannu Jännes, Magnus Hellström, Robin Hemming, Peter Kennerley, Mike Langman, Paul Leader, Petteri Lehikoinen, Antero Lindholm, Vladimir Loskot, Irina Marova, John Martin, Richard Millington, Steve Nuttall, Martyn Pitt, Jelmer Poelstra, Brett Richards, Roger Riddington, Kees Roselaar, Steve Seal, Daria Shipilina, Brian Small, Andy Stoddart, Lars Svensson, Pavel Tomkovich, Chris Turner, Vincent van der Spek, Grahame Walbridge, Andy Warr and Maarten Wielstra. Mark Adams kindly arranged access to the skin collection at NHM, Tring.
Citation: Dean, A. R. 2009 (updates to 2025). 'Siberian Chiffchaff' Phylloscopus collybita tristis: discussion and photo gallery. https://deanar.org.uk/tristis/tristis.htm.
It is intended to keep the contents updated as new information emerges. Thus, any citation should include the date on which the text was consulted.
|
|